Моча образуется в результате обмена растворенными веществами и водой между фильтратом, покидающим проксимальный извитой каналец, и всеми последующими отделами нефрона (табл. 19.7).
Таблица 19.7. Состав плазмы и мочи и изменения концентрации в них растворенных веществ в процессе образования мочи у человека
Способность к образованию гипертонической мочи характерна только для тех позвоночных (птицы и млекопитающие), у которых имеется петля Генле. Концентрация образующейся мочи находится в прямой зависимости от длины петли Генле и толщины слоя мозгового вещества по сравнению с корой. Оба показателя возрастают с увеличением дефицита воды в зоне обитания. Например, у бобра (Castor) — полуводного млекопитающего — имеется тонкий слой мозгового вещества, состоящего из коротких петель Генле, и выделяется большой объем разведенной мочи (600 мосмоль/л), тогда как обитатели пустынь — кенгуровая крыса (Dipodomys) и тушканчик (Dipus) — обладают толстым слоем мозгового вещества с длинными петлями Генле и выделяют небольшое количество сильно концентрированной мочи (6000 и 9000 мосмоль/л соответственно).
Прежде чем подробно обсуждать строение и функцию различных участков петли Генле, необходимо рассмотреть ее функцию в целом. Петля Генле вместе с капиллярами прямых сосудов и собирательной трубкой создает и поддерживает продольный градиент осмотического давления в мозговом веществе по направлению от коркового вещества к сосочку за счет повышения концентраций хлористого натрия и мочевины. Благодаря этому градиенту возможно удаление все большего количества воды путем осмоса из просвета канальца в интерстициальные пространства мозгового вещества, откуда она переходит в прямые сосуды. В конечном счете в соединительной трубке образуется гипертоническая моча.
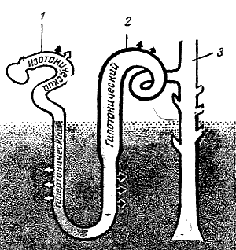
Механизм, с помощью которого петля Генле обеспечивает повышение концентрации хлористого натрия в мозговом веществе по направлению к сосочку, не вполне ясен. Не известна также степень проницаемости нисходящего колена петли Генле для хлористого натрия. Эта неясность не позволяет полностью понять механизм образования градиента, но не изменяет существенно того, что было сказано выше и будет сказано ниже. Движение ионов, мочевины и воды между петлей Генле, прямыми сосудами и собирательной трубкой можно описать следующим образом (см. соответствующие цифры в кружках на рис. 19.26):
Рис. 19.26 Общая схема, показывающая передвижение солей, мочевины и воды, в результате которого создается градиент осмолярности в мозговом веществе. Цифры в прямоугольниках показывают осмолярность (мосмоль/кг), а цифры в кружочках соответствуют пунктам текста. Тоничность разных отделов нефрона указана по отношению к плазме крови за пределами почки
1. Короткий и относительно широкий (30 мкм) верхний сегмент нисходящего колена непроницаем * для солей, мочевины и воды. По этому участку фильтрат переходит из проксимального извитого канальца в более длинный тонкий (12 мкм) сегмент нисходящего колена, свободно пропускающий воду.
* ( Здесь и далее речь идет, конечно, о проницаемости стенок.)
2. Благодаря высокой концентрации хлористого натрия и мочевины в тканевой жидкости мозгового вещества создается высокое осмотическое давление, вода отсасывается из фильтрата и поступает в прямые сосуды.
3. В результате выхода воды из фильтрата его объем уменьшается на 5% и он становится гипертоничным. В верхушке мозгового вещества (в сосочке) нисходящее колено петли Генле изгибается в виде шпильки для волос и переходит в восходящее колено, которое по всей своей длине проницаемо для воды.
4. Нижний участок восходящего колена — тонкий сегмент — проницаем для хлористого натрия и мочевины, и хлористый натрий диффундирует из него, тогда как мочевина, напротив, диффундирует внутрь.
5. В следующем, толстом сегменте восходящего колена эпителий состоит из уплощенных кубовидных клеток с рудиментарной щеточной каемкой и многочисленными митохондриями. В этих клетках осуществляется активный перенос ионов натрия и хлора из фильтрата.
6. Вследствие выхода ионов натрия и хлора из фильтрата повышается осмолярность мозгового вещества, а в дистальные извитые канальцы поступает гипотоничный фильтрат.
Роль петли Генле как противоточного концентрирующего механизма определяют следующие факторы:
1) близкое соседство нисходящего и восходящего колен;
2) проницаемость нисходящего колена для воды;
3) непроницаемость нисходящего колена для растворенных веществ;
4) проницаемость тонкого сегмента восходящего колена для растворенных веществ;
5) наличие механизмов активного транспорта в толстом сегменте восходящего колена.
Осмолярность фильтрата в обоих коленах петли Генле тоже позволяет предположить, что петля действует как противоточный концентрирующий механизм. В такой системе разность концентраций между жидкостями в восходящем и нисходящем коленах на каждом уровне может быть небольшой, но поскольку эта разность существует на большом протяжении (как в петле Генле), она суммируется и конечная концентрация фильтрата у изгиба петли оказывается намного больше, чем у ее концов. Чем длиннее петля, тем больше разность концентраций. Выход хлористого натрия из фильтрата в восходящем колене приводит к повышению его концентрации в тканевой жидкости мозгового вещества, в связи с чем из восходящего колена отсасывается вода. Эта вода сразу поступает в прямые сосуды, благодаря чему в тканевой жидкости и в фильтрате сохраняется высокая концентрация растворенных веществ, как показано на рис. 19.27. В процессе продвижения фильтрата по петле противоточный обмен продолжается. Непрерывная пассивная и активная циркуляция растворенных веществ обеспечивает постоянную высокую концентрацию хлористого натрия в мозговом веществе. Параллельно функционирует другая система-система противоточного обмена в прямых сосудах (см. ниже), и эти два противоточных механизма, работая сообща, обеспечивают градиент осмотической концентрации в мозговом веществе.
Рис. 19.27. Модель противоточного концентрирующего механизма, образуемого канальцами петли Генле. Ионы Na + и Сl — активно транспортируются из толстого сегмента восходящего колена петли в тканевую жидкость, концентрация растворенных веществ в этой жидкости повышается. Вода выходит из просвета нисходящего колена петли. Цифрами указана осмотическая концентрация (мосмоль/кг). Градиент осмолярности между нисходящим и восходящим коленами петли Генле произвольно принят за 200 мосмоль/кг
Узкий нисходящий и более широкий восходящий капилляры прямых сосудов на всем протяжении идут параллельно друг другу и образуют на разных уровнях ветвящиеся петли. Эти капилляры проходят очень близко к канальцам петли Генле, однако прямого переноса веществ из фильтрата петли в прямые сосуды не происходит. Вместо этого растворенные вещества выходят сначала в интерстициальные пространства мозгового вещества, где мочевина и хлористый натрий задерживаются из-за малой скорости кровотока в прямых сосудах, и осмотический градиент тканевой жидкости сохраняется. Клетки стенок прямых сосудов свободно пропускают воду, мочевину и соли, а поскольку эти сосуды идут рядом, они функционируют как система противоточного обмена. При вступлении нисходящего капилляра в мозговое вещество из плазмы крови вследствие прогрессирующего повышения осмотического давления тканевой жидкости выходит путем осмоса вода, а обратно входят путем диффузии хлористый натрий и мочевина (рис. 19.28). В восходящем капилляре происходит обратный процесс: в связи с уменьшением осмотического давления тканевой жидкости вода вновь переходит в плазму, а хлористый натрий и мочевина выходят из нее. Адаптивное значение этого механизма состоит в том, что благодаря ему осмотическая концентрация плазмы, выходящей из почек, остается стабильной независимо от концентрации плазмы, поступающей в них. Наконец, что особенно важно, поскольку все перемещения растворенных веществ и воды происходят пассивно, противоточный обмен в прямых сосудах происходит без всяких затрат энергии.
Рис. 19.28. Пассивное перемещение воды, ионов и мочевины между соседними прямыми сосудами и между этими сосудами и тканевой жидкостью мозгового вещества. Прямые сосуды образуют систему противоточного обмена. Цифры — величины осмолярности (моемо ль/кг)
Дистальный извитой каналец подходит обратно по направлению к мальпигиеву тельцу и весь лежит в корковом веществе. Клетки дистальных канальцев имеют щеточную каемку и содержат много митохондрий, и именно этот отдел нефронов ответствен за тонкую регуляцию водно-солевого баланса и рН крови. Гормональный контроль проницаемости клеток дистального извитого канальца для воды связан с таким же контролем собирательных трубок (оба механизма описаны ниже). Регуляция баланса электролитов и рН описана в разд. 19.7.
Собирательная трубка начинается в корковом веществе от дистального извитого канальца и идет вниз через мозговой слой, где объединяется с несколькими другими собирательными трубками в более крупные протоки, называемые протоками Беллини. Проницаемость стенок собирательной трубки для воды и мочевины регулируется антидиуретическим гормоном (АДГ), и благодаря этой регуляции собирательная трубка участвует вместе с дистальным извитым канальцем в образовании гипертонической или гипотонической мочи в зависимости от потребности организма в воде.
источник
Определяющую роль в работе противоточного механизма играет восходящее колено петли Генле. Его эпителий активно реабсорбирует в окружающее интерстициальное пространство ионы натрия, поддерживая гиперосмолярность интерстициальной жидкости. Транспорт натрия сопряжен с транспортом калия и хлора (Na, К, 2С1-котранспорт) и антипортом с ионами водорода, осуществляется посредством соответствующих переносчиков. Проницаемость для воды здесь очень низкая из-за отсутствия аквапоринов.В нисходящем колене моча из-за всасывания воды становится все более и более гиперосмотичной, т. е. устанавливается осмотическое равновесие с интерстициальной жидкостью. В восходящем колене из-за всасывания натрия моча становится все менее осмотичной и в корковый отдел дистального канальца восходит уже гипотоничная моча. Однако ее количество из-за всасывания воды и солей в петле Генле существенно уменьшилось.
Стенка собирательной трубочки становится проницаемой для воды только в присутствии вазопрессина.На базолатеральных мембранах клеток собирательных трубочек постоянно встроены независимые от вазопрессина аквапорины 3-го и 4-го типов, свободно пропускающие воду в интерстициальное пространство. По мере продвижения мочи по собирательным трубочкам вглубь мозгового вещества вода пассивно уходит в ги-перосмотичный интерстиций и моча становится все более концентрированной.
Под влиянием вазопрессина реализуется еще один важный для концентрирования мочи механизм: активация переносчика, усиливающая выход мочевины из собирательных трубочек, расположенных в глубине мозгового вещества, в окружающий интерстиций. Всасывание воды в верхних отделах собирательных трубочек ведет к нарастанию концентрации мочевины в моче, а в самых нижних их отделах, расположенных в глубине мозгового вещества, вазопрессин повышает проницаемость для мочевины и она пассивно диффундирует в интерстиций, резко повышая его осмотическое давление. Таким образом, интерстиций мозгового вещества становится наиболее высоко осмотичным в области вершины почечных пирамид, где и происходит увеличение всасывания воды из просвета канальцев в интерстиций и концентрирование мочи.
Мочевина интерстициальной жидкости по концентрационному градиенту диффундирует в просвет тонкой восходящей части петли Генле и вновь поступает с током мочи в дистальные канальцы и собирательные трубочки. Так осуществляется кругооборот мочевины в канальцах, сохраняющий высокий уровень ее концентрации в мозговом веществе. Описанные процессы транспорта мочевины осуществляются, в основном, в юкстамедуллярных нефронах, имеющих наиболее длинные петли Генле, спускающиеся глубоко внутрь мозгового вещества почки.
В мозговом веществе почки имеется и другая — сосудистая противоточная система, образованная кровеносными капиллярами. Кровеносная сеть юкстамедуллярных нефронов образует длинные параллельные прямые нисходящие и восходящие капиллярные сосуды, спускающиеся вглубь мозгового вещества. Двигающаяся по нисходящему прямому капилляру кровь постепенно отдает воду в окружающее интерстициальное пространство в силу нарастающего осмотического давления в ткани и, напротив, обогащается натрием и мочевиной, сгущается и замедляет свое движение. В восходящем капиллярном отделе сосуда по мере движения крови в ткани с постепенно снижающимся осмотическим давлением происходят обратные процессы — натрий и мочевина по концентрационному градиенту диффундируют обратно в ткань, а вода всасывается в кровь. Таким образом, сосудистая противоточная система не создает, а лишь способствует поддержанию высокого осмотического давления в глубоких слоях ткани мозгового вещества почек, обеспечивая удаление воды и удержание натрия и мочевины в интерстиции.
Не нашли то, что искали? Воспользуйтесь поиском:
источник
В почках предусмотрен механизм, позволяющий регулировать осмотическое давление плазмы крови за счет концентрирования или, наоборот, разведения мочи. Этот механизм получил название — поворотно-противоточно-множительный механизм, который локализован в петле Генле и собирательных трубках. Именно в этой части нефрона происходит изменение осмотической концентрации, или осмотического давления мочи, благодаря чему осуществляется регулирование объема выводимой жидкости и ее осмотического давления. Принцип работы этого механизма заключается в следующем: когда первичная моча проходит по собирательной трубке и направляется в чашечку, а затем в лоханку, из нее выходит вода, благодаря чему резко уменьшается объем конечной мочи, и возрастает концентрация в моче осмотически активных веществ. Если бы этот процесс не происходил, то ежедневно, помимо обычных 1—1,5 л мочи, выделялось еще 20—24 л мочи. Когда первичная моча входит в начальную часть собирательной трубки, ее осмотическое давление составляет примерно 290—300 мосмоль (милиосмоль — это единица концентрации осмотически активных веществ, или единица осмотического давления, например, раствор 140 ммоль хлористого натрия представляет собой 140 мосмоль натрия + 140 мосмоль хлора = 280 мосмоль, а раствор, содержащий 140 ммоль хлористого натрия и 10 ммоль хлористого калия имеет осмолярность, равную 140 + 140 + 10 +10 = 300 мосмоль/л.). По мере прохождения мочи по собирательной трубке вода покидает трубку и поэтому осмотическое давление (осмолярность мочи) возрастает до 900—1200 мосмоль, т. е. концентрируется в 3—4 раза. Для такого концентрирования необходимо, чтобы в интерстиции по мере приближения трубки к сосочку (по направлению к чашечке) росло осмотическое давление. В этом случае на каждом «этаже» вода будет покидать трубку и моча будет становиться все более концентрированной.
Выход воды из собирательной трубки происходит за счет осмотического давления, которое создается вокруг собирательной трубки в интерстиции. Это давление обусловлено наличием в интерстиции осмотически активных веществ — ионов натрия, хлора, мочевины. При этом необходимым условием должно быть постепенное повышение концентрации осмотически активных веществ по направлению от наружного слоя к внутреннему слою мозговой части почки. Осмотически активные вещества — натрий и хлор — появляются в интерстиции благодаря работе эпителия восходящей части петли Генле, за счет активного транспорта происходит выход ионов натрия и хлора из первичной мочи, идущей по восходящей части петли Генле. Мочевина появляется в интерстиции благодаря выходу из собирательных трубок за счет градиента концентрации между мочой, находящейся в собирательной трубке, и содержимого интерстиции (см. выше, реабсорбция мочевины). Для того, чтобы из восходящей части петли Генле происходила реабсорбция натрия и ионов хлора, в нисходящей части петли Генле осуществляется выход воды в интерстициальное пространство, где ионы натрия и хлора, вышедшие из восходящей части петли Генле создают осмотически активную среду. Рассмотрим пример. В нисходящую часть петли Генле входит первичная моча, осмолярность которой равна 300 мосмоль/л. По мере продвижения мочи к повороту за счет осмотически активной среды в интерстиции вода из этой порции мочи
будет уходить, и постепенно концентрация осмотически активных веществ будет возрастать — 400,500,600 мосмоль, достигая на вершине петли Генле максимальной концентрации, например, 1200 мосмоль. Затем, поднимаясь по восходящей части петли Генле, эпителий которой непроницаем для воды, но проницаем для ионов натрия и хлора, натрий и хлор за счет разности концентрации будут выходить в интерстиций (это и необходимо для конечной концентрации мочи в собирательных трубках), поэтому осмолярность первичной мочи по мере подъема ее по восходящей части петли Генле будет снижаться: 1000,900,800,700 мосмоль и т. д., а на вершине восходящей части петли Генле осмолярность может стать ниже исходной (например, 200,100 мосмоль). Когда моча попадает в собирательную трубку, то за счет высокой проницаемости для воды (но не для натрия и хлора), вода будет покидать собирательную трубку, а осмотическая концентрация (осмолярность) будет возрастать на тем большую величину, чем дальше проходит моча.
Итак, восходящая часть петли Генле создает условия для концентрации мочи как в нисходящей части петли Генле, так и в собирательных трубках.
Проницаемость собирательной трубки для воды зависит от уровня в крови вазопрессина (антидиуретического гормона): чем он выше, тем выше проницаемость для воды, тем меньше организм теряет воды, тем концентрированнее моча. И наоборот, когда уровень АДГ в крови снижается, тогда проницаемость для воды падает, и объем выделяемой мочи возрастает, а концентрация осмотически активных веществ в моче снижается.
Работа такого механизма позволяет организму тонко регулировать осмотическое давление плазмы крови и других жидкостей — при увеличении осмотического давления крови увеличивается реабсорбция воды в собирательных трубках, возрастает объем циркулирующей крови, что приводит к нормализации осмотического давления плазмы. Наоборот, при уменьшении осмотического давления почки в большем объеме, чем обычно, выделяют воду, что приводит к нормализации осмотического давления.
Важная роль в процессах концентрирования мочи принадлежит мозговому кровотоку. Если этот кровоток очень интенсивный, то из интерстиция удаляются осмотически активные вещества — натрий, хлор, мочевина. Тем самым снижается способность концентрировать мочу, возрастает диурез. Вот почему при повышении системного артериального давления вследствие роста интенсивности мозгового кровотока снижается концентрация осмотически активных веществ в мозговом слое почки и повышается диурез.
Для того, чтобы сохранить градиент концентрации осмотически активных веществ в мозговом слое (по направлению к внутренней части мозгового слоя он возрастает), капилляры мозгового слоя почки располагаются в виде петель, параллельно петлям Генле.
Итак, представленные данные позволяют понять принцип работы поворотно-противо-точно-множительной системы: за счет параллельного расположения трубок (нисходящей и восходящей частей петли Генле, собирательной трубки) создается противоток, в результате которого умножаются градиенты концентрации осмотически активных веществ.
источник
Увеличение кровотока в мозговом слое может приводить к снижению концентрирующей способности почек. Некоторые вазодилататоры способны значительно увеличивать кровоснабжение мозгового слоя, таким образом вымывая из него некоторые вещества и уменьшая максимальную концентрирующую способность почек. Значительное повышение артериального давления также способствует увеличению кровотока в мозговом слое почки, причем в значительно большей степени, чем в других отделах, что ведет к вымыванию веществ и уменьшению способности почек концентрировать мочу. Как обсуждалось ранее, максимальная концентрирующая способность почки определяется не только уровнем АДГ, но и осмолярностью межклеточной жидкости мозгового вещества. Даже при максимальном уровне АДГ концентрирующая способность почки будет снижена, если кровоток в мозговом слое возрастет настолько, что уменьшит его осмолярность.
Проксимальный каналец. Около 65% попавших в первичную мочу электролитов реабсорбируется в проксимальном канальце. Однако мембраны канальцев высокопроницаемы и для воды: в то время, когда реабсорбируются растворенные вещества, вода вследствие осмоса диффундирует через стенку канальца. Следовательно, осмолярность жидкости остается такой же, как и в фильтрате клубочка, составляя 300 мосм/л.
Нисходящий отдел петли Генле. Движение мочи по нисходящему отделу петли Генле способствует поглощению воды в мозговом слое. Этот отдел высокопроницаем для воды, но в значительно меньшей степени — для NaCl и мочевины. Следовательно, осмолярность мочи в ее просвете постепенно возрастает, увеличиваясь до тех пор, пока не сравняется с окружающей межклеточной жидкостью, которая при высокой концентрации АДГ составляет 1200 мосм/л. При образовании почками разведенной мочи вследствие низкой концентрации АДГ осмолярность межклеточной жидкости мозгового слоя составит менее 1200 мосм/л; следовательно, моча в просвете нисходящего отдела также становится менее концентрированной. Снижение осмолярности отчасти объясняется меньшей реабсорбцией мочевины из собирательных протоков во внутренний отдел мозгового слоя почки вследствие низкого уровня АДГ, в результате почка выделяет большой объем разведенной мочи.
Тонкий сегмент восходящего отдела петли Генле. Тонкий сегмент восходящего отдела петли Генле практически непроницаем для воды, но позволяет диффундировать небольшому количеству NaCl. Высокая концентрация NaCl в просвете, обусловленная удалением воды в нисходящем отделе петли Генле, способствует небольшой диффузии NaCl в мозговой слой. Итак, моча становится более разведенной, поскольку NaCl покидает просвет, а вода остается. Небольшое количество мочевины, перешедшей из собирательных протоков в мозговой слой также путем диффузии, проникает в восходящий отдел, возвращая таким образом мочевину в канальцевую систему и препятствуя вымыванию из мозгового слоя почки. Повторное использование мочевины представляет собой дополнительный механизм, благодаря которому поддерживается высокая осмолярность мозгового слоя почки.
Толстый сегмент восходящего отдела петли Генле. Данный сегмент нефрона также практически непроницаем для воды, однако здесь происходит активный транспорт значительного количества ионов Na+, Сl-, К+, а также других ионов из жидкости внутри просвета канальцев в мозговой слой почки. Следовательно, моча в толстом восходящем отделе петли Генле становится сильно разведенной, осмолярность ее падает до 100 мосм/л.
Начальный отдел дистального канальца. Начальная часть дистального канальца по свойствам напоминает толстый восходящий отдел петли Генле, поэтому благодаря реабсорбции растворенных веществ здесь происходит дальнейшее разведение мочи, поскольку вода остается в просвете канальца.
85. Регуляция водно-солевого обмена у человека: роль вазопрессина, альдостерона и натрийуретических пептидов.
Водно-солевой обмен
– совокупность процессов поступления воды и солей (электролитов) в организм, их всасывания, распределения во внутренних средах и выделения. Суточное потребление человеком воды составляет около 2,5 л, из них около 1 л он получает с пищей. В организме человека 2/3 общего количества воды приходится на внутриклеточную жидкость и 1/3 – на внеклеточную. Часть внеклеточной воды находится в сосудистом русле (около 5% от массы тела), большая же часть внеклеточной воды находится вне сосудистого русла, это межуточная (интерстициальная), или тканевая, жидкость (около 15% от массы тела). Кроме того, различают свободную воду, воду, удерживаемую коллоидами в виде так называемой воды набухания, т.е. связанную воду, и конституционную (внутримолекулярную) воду, входящую в состав молекул белков, жиров и углеводов и освобождающуюся при их окислении. Разные ткани характеризуются различным соотношением свободной, связанной и конституционной воды. За сутки почками выводится 1–1,4 л воды, кишечником – около 0,2 л; с потом и испарением через кожу человек теряет около 0,5 л, с выдыхаемым воздухом – около 0,4 л.
Системы регуляции водно-солевого обмена обеспечивают поддержание общей концентрации электролитов (натрия, калия, кальция, магния) и ионного состава внутриклеточной и внеклеточной жидкости на одном и том же уровне. В плазме крови человека концентрация ионов поддерживается с высокой степенью постоянства и составляет (в ммоль/л): натрия – 130–156, калия – 3,4–5,3, кальция – 2,3–2,75 (в т.ч. ионизированного, не связанного с белками – 1,13), магния – 0,7–1,2, хлора – 97–108, бикарбонатного иона – 27, сульфатного иона – 1,0, неорганического фосфата – 1–2. По сравнению с плазмой крови и межклеточной жидкостью клетки отличаются более высоким содержанием ионов калия, магния, фосфатов и низкой концентрацией ионов натрия, кальция, хлора и ионов бикарбоната. Различия в солевом составе плазмы крови и тканевой жидкости обусловлены низкой проницаемостью капиллярной стенки для белков. Точная регуляция водно-солевого обмена у здорового человека позволяет поддерживать не только постоянный состав, но и постоянный объем жидкостей тела, сохраняя практически одну и ту же концентрацию осмотически активных веществ и кислотно-щелочное равновесие.
Регуляция водно-солевого обмена осуществляется при участии нескольких физиологических систем. Сигналы, поступающие от специальных неточных рецепторов, реагирующих на изменение концентрации осмотически активных веществ, ионов и объема жидкости передаются в ЦНС, после чего выделение из организма воды и солей и их потребление организмом меняется соответствующим образом. Так, при увеличении концентрации электролитов и уменьшении объема циркулирующей жидкости (гиповолемии) появляется чувство жажды, а при увеличении объема циркулирующей жидкости (гиперволемии) оно уменьшается. Увеличение объема циркулирующей жидкости за счет повышенного содержания воды в крови (гидремия) может быть компенсаторным, возникающим после массивной кровопотери. Гидремия представляет собой один из механизмов восстановления соответствия объема циркулирующей жидкости емкости сосудистого русла. Патологическая гидремия является следствием нарушения водно-солевого обмена, например при почечной недостаточности и др. У здорового человека может развиться кратковременная физиологическая гидремия после приема больших количеств жидкости. Выведение воды и ионов электролитов почками контролируется нервной системой и рядом гормонов. В регуляции водно-солевого обмена участвуют и вырабатываемые в почке физиологически активные вещества – производные витамина D3, ренин, кинины и др.
Содержание натрия и организме регулируется в основном почками под контролем ЦНС через специфические натриорецепторы. реагирующие на изменение содержания натрия в жидкостях тела, а также волюморецепторы и осморецепторы, реагирующие на изменение объема циркулирующей жидкости и осмотического давления внеклеточной жидкости соответственно. Натриевый баланс в организме контролируется и ренин-ангиотензинной системой, альдостероном, натрийуретическими факторами. При уменьшении содержания воды в организме и повышении осмотического давления крови усиливается секреция вазопрессина (антидиуретического гормона), который вызывает увеличение обратною всасывания воды в почечных канальцах. Увеличение задержки натрия почками вызывает альдостерон, а усиление выведения натрия – натрийуретические гормоны, или натрийуретические факторы. К ним относятся атриопептиды, синтезирующиеся в предсердиях и обладающие диуретическим, натрийуретическим действием, а также некоторые простагландины, уабаинподобное вещество, образующееся в головном мозге, и др.
Основным внутриклеточным кучным осмотически активным катионом и одним из важнейших потенциал образующих ионов является калий. Мембранный потенциал покоя, т.е. разность потенциалов между клеточным содержимым и внеклеточной средой, сознается благодаря способности клетки активно с затратой энергии поглощать ионы К+ из внешней среды в обмен на ионы Na+ (так называемый К+, Na+-насос) и вследствие более высокой проницаемости клеточной мембраны для ионов К+ чем для ионов Na+. Из-за высокой проницаемости неточной мембраны для ионов К+ дает небольшие сдвиги в содержании калия в клетках (в норме это величина постоянная) и плазму крови ведут к изменению величины мембранного потенциала и возбудимости нервной и мышечной ткани. На конкурентных взаимодействиях между ионами К+ и Na+, а также К+ и Н+ основано участие калия в поддержании кислотно-щелочного равновесия в организме. Увеличение содержания белка в клетке сопровождается повышенным потреблением ею ионов К+. Регуляция обмена калия в организме осуществляется ц.н.с. при участии ряда гормонов. Важную роль в обмене калия играют кортикостероиды, в частности альдостерон, и инсулин.
При дефиците калия в организме страдают клетки, а затем наступает гипокалиемия. При нарушении функции почек может развиваться гиперкалиемия, сопровождаемая тяжелым расстройством функций клеток и кислотно-щелочного состояния. Нередко гиперкалиемия сочетается с гипокальциемией, гипермагниемией и гиперазотемией.
Состояние водно-солевого обмена в значительной степени определяет содержание ионов Cl – во внеклеточной жидкости. Из организма ионы хлора выводятся в основном с мочой. Количество экскретируемого хлорида натрия зависит от режима питания, активной реабсорбции натрия, состояния канальцевого аппарата почек, кислотно-щелочного состояния и др. Обмен хлоридов тесно связан с обменом воды: уменьшение отеков, рассасывание транссудата, многократная рвота, повышенное потоотделение и др. сопровождаются увеличением выведения ионов хлора из организма. Некоторые диуретики с салуретическим действием угнетают реабсорбцию натрия в почечных канальцах и вызывают значительное увеличение экскреции хлора с мочой. Многие заболевания сопровождаются потерей хлора. Если его концентрация в сыворотке крови резко снижается (при холере, острой кишечной непроходимости и др.), прогноз заболевания ухудшается. Гиперхлоремию наблюдают при избыточном потреблении поваренной соли, остром гломерулонефрите, нарушении проходимости мочевых путей, хронической недостаточности кровообращения, гипоталамо-гипофизарной недостаточности, длительной гипервентиляции легких и др.
При ряде физиологических и патологических состояний часто бывает необходимо определить объем циркулирующей жидкости. С этой целью в кровь вводят специальные вещества (например, краситель синий Эванса или меченный альбумин). Зная количество вещества, введенного в кровоток, и определив через некоторое время его концентрацию в крови, рассчитывают объем циркулирующей жидкости. Содержание внеклеточной жидкости определяют с помощью веществ, не проникающих внутрь клеток. Общий объем воды в организме измеряют по распределению «тяжелой» воды D2O, воды, меченной тритием [рН] 2О (ТНО), или антипирина. Вода, в состав которой входит тритий или дейтерий, равномерно смешивается со всей водой, содержащейся в теле. Объем внутриклеточной воды равен разности между общим объемом воды и объемом внеклеточной жидкости.
Осмоляльность плазмы крови и внеклеточной жидкости определяется главным образом натрием, поскольку натрий является основным внеклеточным катионом, и 85% эффективного осмотического давления зависит от натрия с сопутствующими анионами. На долю остальных осмотически активных веществ приходится примерно 15%, и регуляция осмоляльности жидкостей внутренней среды фактически сводится к поддержанию постоянства соотношения воды и натрия. Экскреция воды почкой регулируется антидиуретическим гормоном нейрогипофиза (АДГ) и в конечном итоге определяется теми факторами, которые влияют на скорость синтеза и секреции АДГ и его эффект в почке.
Сенсорный механизм антидиуретической системы представлен осморецепторами с высокой чувствительностью к отклонению осмоляльности плазмы крови. После открытия английским физиологом Е. Вернеем осмочувствительных элементов в гипоталамусе дальнейший прогресс в изучении локализации и функции центральных осморецепторов был обусловлен развитием электрофизиологических исследований и радиоиммунного способа определения концентрации АДГ. В опытах на различных животных было установлено, что при введении через катетер в сонную артерию или непосредственно в мозг через микроэлектрод 2%-ного раствора хлорида натрия увеличивается активность отдельных нейронов, расположенных в зоне III желудочка. Такие нейроны располагались в области супраоптического и паравентрикулярного ядер, то есть скопления крупноклеточных нейронов над перекрестом зрительного тракта и около стенки III желудочка, в которых осуществляется синтез АДГ – стимулятора реабсорбции воды в почке. Осморецепторы мозга сигнализируют об отклонениях от нормального уровня осмоляльности крови, притекающей к мозгу.
Однако в системе, регулирующей баланс воды, мониторинг осмотического равновесия обеспечивается не только осморецепторами мозга. Идея о том, что осморецепторы могут локализоваться не только в мозгу, но и в других тканях, прежде всего в печени, куда притекает кровь от желудочно-кишечного тракта, принадлежит А.Г. Гинецинскому. Применение тонких сосудистых зондов, с помощью которых в кровоток вводили гипертонические растворы в таком количестве, что осмоляльность крови отклонялась всего на 2 – 5% только в исследуемом органе, но не изменялась в общем кровотоке, позволило выявить осмочувствительные элементы практически во всех висцеральных органах и в скелетных мышцах конечностей. Наиболее активной оказалось осморецептивное поле печени. Электрофизиологическим методом было зарегистрировано повышение импульсной активности в тончайших нервных волокнах, идущих от печени в составе блуждающего нерва и задних корешков мозга в центральную нервную систему Антидиуретическая реакция, то есть резкое ограничение выделения жидкости почками в ответ на осмотическое раздражение различных зон, имеет рефлекторную природу, так как разрыв нервных связей, идущих в мозг к гипоталамическим ядрам, или перерезка ножки гипофиза, осуществляющей нервную связь гипоталамических ядер с задней долей гипофиза, где АДГ выделяется в кровь, предотвращают развитие антидиуреза.
Антидиуретический гормон – основной гормон, регулирующий реабсорбцию воды в почечных канальцах. Второе название этого гормона – вазопрессин, так как он принимает участие в регуляции сосудистого тонуса, стимулируя сокращение гладких мышц артерий и повышая кровяное давление. В последние годы установлено, что эффекты АДГ-вазопрессина весьма разнообразны вплоть до участия в механизмах памяти. Однако, несмотря на множественность действия этого гормона, у млекопитающих и человека наиболее ярко проявляется его роль в регуляции осмоляльности жидкостей внутренней среды. За последние десятилетия были изучены не только структура и свойства этого пептидного гормона, состоящего из 9 аминокислот, но также структура гена, кодирующего этот гормон и пути его биосинтеза.
АДГ синтезируется в нейросекреторных клетках гипоталамуса (супраоптическом и паравентрикулярном ядрах), по отросткам этих клеток с током аксоплазмы перемещается в заднюю долю гипофиза и из окончаний нервных отростков попадает в кровь. При раздражении осморецепторов происходят рефлекторная активация нейронов и выбрасывание в кровь АДГ. В обычных условиях АДГ присутствует в плазме крови в очень низкой концентрации (10 пикомолей на 1 л). Но почки чрезвычайно чувствительны к АДГ, и требуются лишь небольшие изменения скорости секреции вазопрессина (2–5 пикограмм в минуту на 1 кг массы тела) для проявления его эффекта. С током крови АДГ достигает клеток конечных отделов почечных канальцев и взаимодействует с белком-рецептором, встроенным в мембрану (типа V2). В других клетках-мишенях действует рецептор V1 и используются иные внутриклеточные механизмы передачи гормонального сигнала. V2-рецептор сопряжен с ферментом аденилатциклазой через особую ГТФ-зависимую единицу. Это приводит к активации образования вторичного посредника – циклического аденозинмонофосфата (цАМФ), к активации специфического фермента протеинкиназы, и завершается цепь встраиванием в мембрану клетки, обращенную в просвет канальца, особого белка аквапорина. Аквапорин образует в липидной гидрофобной мембране поры, проницаемые для воды, и в результате вода диффундирует в интерстициальное пространство, окружающее собирательные трубки. При этом проявляется антидиурез – резкое ограничение выведения воды из организма.
Уровень секреции АДГ находится под непосредственным влиянием изменений не только осмоляльности плазмы крови, но и объема циркулирующей крови и уровня кровяного давления. Впервые в 1956 году Дж. Генри и О. Гауэр в экспериментах на собаках установили, что повышение давления в левом предсердии приводит к увеличению выделения воды почками. Эти данные, а также последующие эксперименты послужили основой для заключения, что рецепторы растяжения левого предсердия, реагирующие на объем притекающей к сердцу крови (волюморецепторы), играют важную роль в секреции АДГ, вызывая торможение нейронов супраоптического ядра (рис. 2). В дальнейшем изучение реакции человека на изменение объема крови в области грудной клетки в результате усиления ее притока от нижних конечностей привлекло особое внимание в связи с развитием космической биологии и медицины. Переход от земной гравитации к невесомости приводит к устранению действия силы тяжести на кровь, находящуюся в нижних конечностях и в брюшной полости; она перераспределяется в вышележащие части тела, переполняя сосуды грудной полости и головы. Это приводит к растяжению предсердий, рефлекторному подавлению секреции АДГ и адекватной физиологической реакции – усилению почечной экскреции жидкости, приводящей к нормализации уровня кровяного давления. Напротив, при возвращении из космического полета недонаполнение сосудов грудной полости приводит к стимуляции секреции АДГ и усилению реабсорбции воды почкой. Нормализация уровня гормона в плазме крови происходила у космонавтов в течение недели и зависела от длительности полетов. Моделирование невесомости у испытуемых при погружении в воду до уровня шеи, при так называемой иммерсии, дало возможность проследить в динамике изменение объема крови и концентрации АДГ в крови. Данные, полученные при исследовании космонавтов и испытуемых, полностью совпали.
Взаимодействие осмотических и объемных стимулов обеспечивает адекватные для ситуации изменения экскреции воды почкой. В нормальных условиях главной детерминантой секреции АДГ является осмотическая концентрация внеклеточной жидкости. При небольших изменениях объема крови и смещениях осмоляльности в первую очередь выравнивается осмотическая концентрация крови. При значительных изменениях объема внутрисосудистой жидкости независимо от направленности осмоляльности крови соподчиненность систем осмо- и волюморегуляции изменяется и включаются прежде всего механизмы, способствующие восстановлению объема.
источник
Противоточно-поворотная система почки. Роль петли Генле в создании кортико-медуллярного осмотического градиента и образовании конечной мочи.
Кора надпочечников человека синтезирует 3 основных класса стероидных гормонов, которые обладают широким спектром физиологических функций. Они включают глюкокортикоиды, минералокортикоиды и адренальные андрогены. Эти гормоны образуются в различных слоях надпочечников из холестерола липопротеинов низкой плотности или ацетил коэнзима А, или эфиров холестерина из внутриклеточных депо. В клубочковом слое коры надпочечников синтезируются гормоны, участвующие в регуляции обмена натрия и воды (альдостерон),
Петля Генле — это часть канальца, погружающаяся или “изгибающаяся” из коркового слоя в мозговой (нисходящее колено), и затем возвращающаяся в кору почек (восходящая колено). Именно в этой части канальца моча при необходимости концентрируется. Это возможно благодаря высокой концентрации веществ в интерстиции мозгового слоя, которая поддерживается за счет наличия “противоточно-поворотной системы”. Противоточно-поворотная система поддерживает высокий осмотический градиент интерстиция мозгового слоя, что позволяет почкам концентрировать мочу. Петля Генле — это противоточно-поворотный множитель, а vasa recta — это противоточно-поворотный обменник, механизм которого описан ниже.
Противоточный механизм состоит в том, что движение канальцевой жидкости в нисходящем и восходящем отделах петли Генле происходит в противоположном направлении, так же как и в венозном (восходящем) и артериальном (нисходящем) отделах прямых сосудов мозгового вещества. Поворотный механизм осуществляется в самом колене петли Генле, где движение канальцевой жидкости получает обратное направление. Концентрационный множительный эффект этой системы обусловлен нарастанием осмотического давления в интерстициальной ткани в направлении от пограничной зоны, где осмотическое давление составляет 280-300 мосмоль/л, к вершине пирамид, где оно достигает 1200-1500 мосмоль/л. В результате создается так называемый вертикальный концентрационный градиент, под влиянием которого происходит реабсорбция воды из канальцев в интерстициальную ткань по всему нисходящему отделу петли Генле, что приводит к нарастанию осмотической концентрации канальцевой жидкости от начала нисходящего отдела петли Генле до его поворота в восходящий отдел.
Функции различных частей петли Генле.
A. Нисходящее колено петли Генле относительно непроницаемо для растворенных веществ и хорошо проницаемо для воды, перемещаемой из канальца по осмотическому градиенту: жидкость в канальце становится гиперосмолярной.
B. Тонкий сегмент восходящего колена петли Генле практически непроницаем для воды, но в то же время проницаем для растворенных веществ, особенно ионов натрия и хлора, которые перемещаются по концентрационному градиенту из просвета канальца, жидкость в котором вначале становится изотоничной, а затем гипотоничной по мере выхода из нее ионов. Мочевина, абсорбировавшаяся в интерстиций мозгового слоя почки из собирательной трубки, диффундирует в восходящее колено. Это поддерживает концентрацию мочевины в интерстиции мозгового слоя, играя важную роль в процессе концентрации мочи.
C. Толстый сегмент восходящего колена петли Генле и начальный отдел дистального канальца непроницаемы для воды. Однако здесь происходит активный транспорт ионов натрия и хлора из просвета канальца, вследствие чего жидкость этого отдела канальца становится крайне гипотоничной.
Собирательные трубочки находятся между многочисленными петлями Генле и идут параллельно им. Под действием АДГ их стенки становятся проницаемыми для воды. Поскольку концентрация солей в петле Генле очень высока, а вода имеет тенденцию следовать за солями, она фактически вытягивается из собирательных трубочек, оставляя раствор с высокой концентрацией солей, мочевины и других растворенных веществ. Этот раствор и есть конечная моча. Если АДГ в крови отсутствует, то собирательные трубочки остаются малопроницаемыми для воды, вода из них не выходит, объем мочи остается большим и она оказывается разведенной.
Осмотическое концентрирование и разведение мочи по принципу ППМС осуществляется следующим образом. В проксимальном отделе канальца вода и осмотически активные вещества (главным образом натрий и мочевина) реабсорбируются в таких соотношениях, что осмолярность канальцевой жидкости остается равной (изоосмотична) осмолярности плазмы крови (280-300 мосмоль/л). Далее канальцевая жидкость поступает в просвет нисходящего отдела петли Генле, стенка которого проницаема для воды и непроницаема для осмотически активных веществ (рис. 11). По мере продвижения канальцевой жидкости по нисходящему отделу петли Генле в направлении от коркового по мозговому веществу почек до вершин пирамид вследствие нарастания осмотического давления в мозговом веществе и при наличии осмотического градиента все больше воды уходит из просвета канальца в интерстиций. Количество канальцевой жидкости постепенно уменьшается, а осмолярность ее возрастает, достигая у места поворота нисходящего отдела петли Генле в восходящий 1200-1500 мосмоль/л. По восходящему тонкому отделу петли Генле канальцевая жидкость движется в обратном направлении (от мозгового вещества почки к корковому) и, следовательно, из зоны наиболее высокого осмотического давления в зону с постепенно уменьшающимся осмотическим давлением в интерстициальной ткани. Поскольку стенка восходящего тонкого отдела петли Генле проницаема для воды, а концентрация осмотически активных веществ в ее просвете выше, чем в окружающем интерстиций, то вода из интерстициальной ткани поступает в просвет этого отдела петли, повинуясь осмотическому градиенту. Достигнув восходящего толстого отдела петли Генле, движение воды внутрь канальца прекращается, так как стенка этого отдела непроницаема для воды и проницаема для натрия. Здесь активно реабсорбируется натрий, но без осмотически эквивалентного количества воды. Это приводит к повышению концентрации натрия и обусловливает образование осмотического градиента в наружной зоне мозгового вещества почки, благодаря чему и осуществляется реабсорбция воды из нисходящего тонкого отдела петли Генле в окружающую интерстициальную ткань. Поскольку в восходящем толстом отделе петли Генле активно реабсорбируется натрий и не реабсорбируется вода, концентрация осмотически активных веществ в просвете этого отдела петли снижается и в дистальный извитый каналец всегда (и при недостатке и при избытке воды в организме) поступает гипотоническая жидкость с концентрацией осмотически активных веществ ниже 200 мосмоль/л. Далее процесс образования мочи идет следующим образом. При недостатке в организме воды (антидиурез) усиливается секреция антидиуретического гормона (АДГ), под влиянием которого повышается проницаемость для воды стенки дистальных отделов канальцев и собирательных трубок, и вода по осмотическому градиенту уходит из просвета канальцев и собирательных трубок в интерстициальную ткань. По мере прохождения канальцевой жидкости по собирательным трубкам в мозговом веществе почки в направлении к вершинам пирамид продолжается дальнейшее сгущение мочи в результате обильной реабсорбции воды. В итоге выделяется мало мочи с высоким содержанием в ней осмотически активных веществ, которое соответствует концентрации этих веществ в интерстициальной ткани мозгового вещества почки на уровне вершины пирамид, т. е. 1200-1500 мосмоль/л. При избытке воды в организме (водный диурез) секреция АДГ прекращается либо резко снижается; в итоге проницаемость стенок дистальных отделов канальцев и собирательных трубок уменьшается либо стенки этих отделов становятся вообще непроницаемыми для воды, реабсорбция которой резко снижается (рис. 12). Натрий же продолжает реабсорбироваться из просвета дистальных отделов канальцев и собирательных трубок в интерстициальную ткань. В результате выделяется много гипоосмолярной мочи, т. е. мочи с низкой концентрацией осмотически активных веществ (около 400-500 мосмоль/л).
5. Рефлекторная регуляция объема крови при гиперволюмии.
Нарушение объема крови проявляется в виде гиповолемии и гиперволемии — уменьшении или увеличении объема крови по сравнению с нормой (нормоволемией), составляющей 6—8% от массы тела, или 65—80 мл крови на 1 кг массы тела. В свою очередь гипо- и гиперволемия подразделяются на простую, полицитемическую и олигоцитемическую в зависимости от того, сохраняется ли при этом нормальное соотношение плазмы и клеток крови (36 — 48% объема крови приходится на долю форменных элементов, 52—64% — на долю плазмы) или же изменяется в сторону преобладания клеток (полицитемическая форма) или плазмы (олигоцитемическая форма). Кроме того, к нарушениям объема крови относят изменения объемного соотношения между форменными элементами и плазмой при нормальном общем объеме крови — олиго- и полицитемическую нормоволемию (гемодилюция и гемоконцентрация). Показателем объемного соотношения является гематокритное число, выражающее содержание форменных элементов (преимущественно эритроцитов) в общем объеме крови (36—48 об.% в норме).
Этиология. Гиповолемия простая (уменьшение объема крови без изменения гематокритного числа) возникает сразу после острой кровопотери и сохраняется до тех пор, пока жидкость не перейдет из тканей в кровь.
Гиповолемия олигоцитемическая (уменьшение объема крови с преимущественным уменьшением в ней клеток — эритроцитов) наблюдается после острой кровопотери, когда компенсаторное поступление крови из депо и тканевой жидкости в кровеносное русло не восстанавливает объем и состав крови.
Гиповолемия полицитемическая (уменьшение объема крови вследствие уменьшения объема плазмы при относительном увеличении содержания эритроцитов) развивается при обезвоживании организма (понос, рвота, усиленное потоотделение, гипервентиляция). При шоке кровь депонируется в расширенных сосудах брюшной полости, что ведет к снижению объема циркулирующей крови, а выход жидкости в ткани при повышении проницаемости сосудистой стенки обусловливает сгущение крови и возникновение полицитемической гиповолемии.
Гиперволемия простая (увеличение объема крови при сохранении нормального соотношения между эритроцитами и плазмой) возникает сразу же после переливания большого количества крови. Однако вскоре жидкость покидает кровеносное русло, а эритроциты остаются, что ведет к сгущению крови. Простая гиперволемия при усиленной физической работе обусловлена поступлением в общий кровоток крови из депо.
Гиперволемия олигоцитемическая (увеличение объема крови за счет плазмы) развивается при задержке воды в организме в связи с заболеванием почек, при введении кровезаменителей. В эксперименте она моделируется путем внутривенного введения животным изотонического раствора натрия хлорида.
Гиперволемия полицитемическая (увеличение объема крови за счет нарастания количества эритроцитов) наблюдается при понижении атмосферного давления, а также при различных заболеваниях, связанных с кислородным голоданием (порок сердца, эмфизема), и рассматривается как компенсаторное явление. При эритремии полицитемическая гиперволемия является следствием опухолевого разрастания клеток костного мозга.
Нормоволемия олигоцитемическая возникает при анемии вследствие кровопотери (объем крови нормализовался за счет тканевой жидкости, а количество эритроцитов еще не восстановилось), гемолиза эритроцитов, нарушения гемопоэза.
Нормоволемия полицитемическая наблюдается при переливании небольших количеств эритроцитарной массы.
Патогенез. Гиповолемия сопровождается нарушением транспортной функции крови и связанных с ней дыхательной, трофической, экскреторной, защитной, регуляторной (гуморальная регуляция, терморегуляция) функций крови, что в той или иной мере отражается на гомеостазе.
Гиперволемия обусловливает повышение нагрузки на сердце, особенно при одновременном возрастании гематокритного числа (полицитемическая гиперволемия), когда увеличивается вязкость крови (внутреннее трение), повышается склонность к образованию тромбов и могут возникнуть нарушения кровообращения в некоторых органах.
Патогенез нарушений, развивающихся при олигоцитемической нормоволемии, следует прежде всего связать со снижением дыхательной функции крови и развитием гипоксии.
Помимо осмо- и натриорецепторов, уровень секреции АДГ определяет активность волюморецепторов, воспринимающих изменение объема внутрисосудистой и внеклеточной жидкости. Ведущее значение в регуляции секреции АДГ имеют рецепторы, которые реагируют на изменение напряжения сосудистой стенки в области низкого давления. Прежде всего это рецепторы левого предсердия, импульсы от которых передаются в ЦНС по афферентным волокнам блуждающего нерва. При увеличении кровенаполнения левого предсердия активируются волюморецепторы и угнетается секреция АДГ, что вызывает усиление мочеотделения. Поскольку активация волюморецепторов в отличие от осморецепторов обусловлена увеличением объема жидкости, т. е. возросшим содержанием в организме воды и солей натрия, возбуждение волюморецепторов приводит к увеличению экскреции почкой не только воды, но и натрия. Эти процессы связаны с секрецией натрийуретического гормона, уменьшением секреции ренина, ангиотензина, альдостерона, при этом снижается тонус симпатической нервной системы, в результате уменьшается реабсорбция натрия и возрастают натрийурез и мочеотделение. В конечном счете восстанавливается объем крови и внеклеточной жидкости.
источник
Сущность работы поворотно-противоточной cистемы состоит в том, что два колена петли — нисходящее и восходящее, тесно соприкасаясь друг с другом, функционируют сопряженно как единый механизм ( рис. 103 ). Эпителий нисходящего (проксимального) отдела петли пропускает воду, но не пропускает ионы натрия. Эпителий же восходящего (дистального) отдела петли обладает способностью активно реабсорбировать ионы натрия, т. е. переводить их из канальцевой мочи в тканевую жидкость почки, но одновременно он не пропускает воду из просвета канальцев в тканевую жидкость.
При прохождении мочи через нисходящие (проксимальные) отделы петли Генле моча постепенно сгущается вследствие перехода воды в тканевую жидкость. Переход воды — пассивный процесс, обусловленный тем, что эпителий расположенного рядом восходящего (дистального) отдела петли активно реабсорбирует ионы натрия, т. е. переводит их из просвета канальца в тканевую (интерстициальную) жидкость; перешедшие в тканевую жидкость ионы натрия притягивают туда молекулы воды, но не из дистального, а из проксимального канальца.
 |