Осмотическое концентрирование и разведение мочи — процесс, обеспечивающий постоянство концентрации осмотически активных веществ в крови. Процесс обеспечивается функцией петли нефрона и собирательных трубочек как «противоточного умножителя». Принцип противотока — движение первичной мочи по коленам петли нефрона и собирательным трубочкам навстречу друг другу — создает возможность уравновешивания концентрации осмотически активных веществ в моче и интерстиции почек посредством пассивного движения воды по осмотическому градиенту.
Принцип «умножителя» обеспечивается благодаря активному транспорту хлорида натрия в интерстиции из толстого отдела восходящего колена петли нефрона, который непроницаем для воды. Этот процесс создает высокий осмотический градиент в наружной зоне мозгового вещества почки и разницу в осмотической концентрации в просвете нисходящего и восходящего колена петли нефрона.
Активный транспорт натрия и хлора обеспечивает увеличение осмотической концентрации по направлению к сосочку почки в наружной зоне мозгового вещества почки. В зоне внутреннего мозгового вещества осмотическая концентрация интерстиция дополнительно возрастает за счет транспорта мочевины — внутрипочечный кругооборот мочевины. Способствуют созданию и поддержанию высокого осмотического градиента интерстиция артериальные и венозные прямые кровеносные сосуды, кровоснабжающие мозговое вещество почки, которые повторяют ход колен петли нефрона.
Осмотическая концентрация первичной мочи (ультрафильтрат плазмы), поступившей в нисходящий отдел петли нефрона, равнозначна плазменной 280—300 мосмль/кг Н20. По ходу продвижения по нисходящему колену петли нефрона, стенка которого проницаема для воды, последняя по осмотическому градиенту переходит из просвета петли в окружающую межуточную ткань, и осмотическая концентрация мочи в нисходящем колене петли, уравновешиваясь с осмотической концентрацией интерстиция, постепенно возрастает, достигая максимума (1400—1450 мосмоль/кг Н20) в месте перегиба петли в зоне сосочка почки. В восходящем колене петли нефрона по направлению к корковому веществу почки осмотическая концентрация мочи постепенно уменьшается и в зоне толстого восходящего отдела петли нефрона составляет 200—250 мосмоль/кг Н20. С такой осмотической концентрацией моча поступает в дистальный каналец нефрона и далее в собирательные трубочки. Окончательная осмоляльность мочи зависит от степени проницаемости собирательных трубочек для воды, которая определяется секрецией антидиуретического гормона (АДГ). В то же время уровень секреции АДГ зависит от степени гидратации организма. При избытке воды в организме секреция АДГ тормозится, собирательные трубочки становятся непроницаемыми для воды, и в результате почки выделяют большой объем гипотонической мочи с низкой концентрацией солей. При дефиците воды в организме секреция АДГ повышена, собирательные трубочки проницаемы для воды; по осмотическому градиенту вода из собирательных трубочек переходит в гиперосмолярный интерстиции, и осмотическая концентрация протекающей по собирательным трубочкам мочи уравновешивается с осмотической концентрацией интерстиция. В результате выделяется небольшой объем мочи с высокой осмотической концентрацией. Максимальные значения осмоляльности мочи достигают максимальных значений осмотической концентрации интерстиция, составляя 1450 мосмоль/кг Н20.
Осмолярность и осмоляльность представляют собой общую концентрацию растворенных частиц в 1 л раствора (осмолярность) или в 1 кг воды (осмоляльность). Осмоляльность крови в значительной степени зависит от концентрации ионов натрия и хлора, в меньшей степени глюкозы и мочевины.
Чаще развивается как осложнение инфекционного заболевания: эпидемического паротита, гриппа, пневмонии, тифа, туберкулеза, бруцеллеза, травмы. Инфекция распространяется гематогенно или лимфогенно.
Опухоли лоханки встречаются сравнительно редко, в возрасте 40—60 лет, чаще у мужчин. Выделяют доброкачественные (папиллома, ангиома) и злокачественные (папиллярный рак, плоскоклеточный рак, слизисто-железистый рак, саркома) опухоли. Метастазы опухоли по лимфатическим сосудам подслизистого слоя в мочеточник и мочевой пузырь.
источник
Почки выделяют большие объемы гипотонической мочи с низкой концентрацией солей. Мочеотделение может достигать 15 % от объема жидкости, профильтровавшейся в клубочках, т.е. возрастает до 18 мл*мин-1*1,73 м-2 поверхности тела, а осмоляльность мочи снижается до 70—50 мосмоль*кг-1 H2O.
В условиях дефицита жидкости в организме, при повышении осмоляльности крови наступает секреция АДГ, и почка осуществляет осмотическое концентрирование мочи. Мочеотделение уменьшается до 0,5 мл*мин-1*1,73 м-2 поверхности тела, осмоляльность мочи у человека может возрастать до 1200—1400 мосмоль*кг-1 H2O.
Сущность этого процесса в общих чертах может быть представлена следующим образом. После реабсорбции 65 % профильтровавшейся жидкости в проксимальном канальце в тонкий нисходящий отдел петли Генле поступает изотоничная плазме крови жидкость. Содержимое тонкого отдела петли становится более концентрированным, так как из его просвета в межклеточное вещество по осмотическому градиенту всасывается вода, а осмотически активные вещества остаются внутри канальца. Чем дальше от коры по продольной оси мозгового вещества находится жидкость в нисходящем колене петли Генле, тем выше ее осмолярная концентрация. Таким образом, в каждых соседних участках нисходящего отдела петли наблюдается лишь небольшое нарастание осмотического давления, но по длиннику мозгового слоя осмотическая концентрация в почке человека увеличивается, постепенно повышаясь от 300 до 1200—1500 мосмоль*кг-1 H2O.
На вершине петли Генле не только в несколько раз возрастает осмотическая концентрация канальцевой жидкости, но и значительно уменьшается ее объем. При дальнейшем движении жидкость проходит по непроницаемому для воды восходящему отделу петли Генле и дистальному канальцу, в которых продолжается реабсорбция ионов хлора и натрия, вода же остается в их просвете. В собирательных трубках коры почки под влиянием АДГ возрастает проницаемость стенки для воды, она начинает реабсорбироваться по осмотическому градиенту. При протекании мочи по собирательным трубкам в мозговом веществе продолжается концентрирование мочи. Поскольку все выше становится осмолярность окружающего собирательные трубки интерстиция, то из просвета всасывается вода, в конечном счете образуется и выделяется гиперосмотическая моча, в которой осмотическое давление так же велико, как в интерстициальной жидкости на вершине почечной пирамиды.
В механизме осмотического концентрирования мочи особую роль играет мочевина. В отличие от наружной зоны мозгового вещества, где повышение осмолярности обусловлено главным образом накоплением солей натрия, во внутреннем мозговом веществе наряду с солями натрия важное значение имеет мочевина. В мозговом веществе почки функционирует специальная система, обеспечивающая кругооборот мочевины и ее удержание в почке. Центральную роль в этом процессе играет неодинаковая проницаемость различных частей канальцев для мочевины и особенно способность АДГ увеличивать проницаемость для мочевины стенки тех участков собирательных трубок, которые расположены во внутреннем мозговом веществе почки. Вследствие реабсорбции воды в вышележащих отделах собирательных трубок в их просвете повышается концентрация мочевины, однако она не может выйти из канальца. Когда же моча достигает того участка собирательных трубок, где АДГ увеличивает проницаемость и для воды, и для мочевины (зона внутреннего слоя мозгового вещества), большие количества обоих веществ реабсорбируются в мозговое вещество. Мочевина увеличивает осмолярную концентрацию в интерстиции мозгового вещества, создавая условия для реабсорбции дополнительных количеств воды. Войдя в ткань мозгового вещества, мочевина диффундирует по межклеточному веществу, проникает в просвет тонкого восходящего отдела петли Генле и движется по канальцу. Общее количество входящей в просвет нефрона мочевины настолько значительно, что при антидиурезе в дистальный извитой каналец поступает больше мочевины, чем профильтровалось в клубочках. Мочевина снова реабсорбируется под влиянием АДГ в собирательных трубках, что и обеспечивает непрестанный ее кругооборот в почке и объясняет важную роль мочевины в процессе осмотического концентрирования мочи.
Эффективность осмотического концентрирования мочи зависит от многих причин. Важное значение имеет соотношение между количеством нефронов с длинной и короткой петлями Генле. Юкстамедуллярные нефроны, петли Генле которых глубоко спускаются в мозговое вещество почки, обеспечивают его высокую осмоляльность и благодаря этому наибольшее осмотическое концентрирование мочи. Очевидно, что степень поражения тех или иных групп нефронов в условиях патологии определяет и дефект процесса осмотического концентрирования мочи.
Увеличение скорости движения жидкости по прямым кровеносным сосудам способствует вымыванию осмотически активных веществ из мозгового вещества почки, а ускорение потока мочи по собирательным трубкам снижает уровень всасывания мочевины, что также ухудшает работу концентрирующего механизма. К этому же приводит питание малобелковой пищей, так как меньше образуется мочевины и не удается достичь высокого осмотического градиента в мозговом веществе почки. В этой же зоне почки образуются простагландины, некоторые из них стимулируют движение крови по прямым сосудам мозгового вещества, а также снижают действие АДГ на проницаемость для воды клеток собирательных трубок.
Оценка состояния осморегулирующей функции почки имеет важное значение для клиницистов. Осмотическое концентрирование мочи требует участия почти всех элементов почки — сосудистой системы, почечных канальцев, интерстиция. Поэтому изменение осморегулирующей функции почки может зависеть от многих причин, их выяснение будет способствовать постановке диагноза. Роль этой функции почки была известна клиницистам еще в начале нашего столетия, когда были предложены тесты на разведение и концентрирование мочи; до наших дней сохранили значение пробы Фольгарда (F.Volhard) и Зимницкого. Нарушение процесса осмотического концентрирования, выделение мочи с низкой относительной плотностью после водной депривации могут быть обусловлены изменением работы любого элемента, описанного выше. Вероятно, для клинициста целесообразно не только рассмотреть данные физиологических исследований, показывающих значение каждого из компонентов системы концентрирования мочи, но и проанализировать особенности работы почки при таких патологических состояниях, которые позволяют глубже оценить роль основных элементов концентрирования мочи и могут быть полезны при выяснении причин полиурии и нарушения концентрационной способности почки. Изменение способности почки к осмотическому разведению и концентрированию мочи может быть обусловлено экстраренальными факторами либо процессами, происходящими в почке.
Описаны случаи нарушения функции осморецепторов, когда почки при сухоядении не концентрировали мочу; это было обусловлено не нарушением функции пептидергических нейронов, как при истинном несахарном диабете, а тем, что нейрогипофиз сохранял способность секретировать АДГ только после инъекции никотина. Полиурия при нормальном состоянии осморецепторов может быть обусловлена повышенной возбудимостью центра жажды, вызывающей увеличенное потребление воды при более низкой осмоляльности плазмы крови, чем у здорового человека. В этих случаях осмоляльность крови ниже, чем в норме (269 мосмоль*кг-1 H2O), в то время как при несахарном диабете она выше, чем в норме (280— 295 мосмоль*кг-1 H2O), и составляет более 295 мосмоль*кг-1 H2O. Во всех этих случаях сохраняется нормальная реакция почки на инъекцию препаратов гормонов задней доли гипофиза или синтетического вазопрессина, что свидетельствует о сохранности осморегулирующей функции почки.
Значительное снижение клубочковой фильтрации и почечного кровотока независимо от вызвавшей их причины приводит к уменьшению концентрационной функции почки из-за недостаточной доставки осмотически активных веществ и нарушения нормального движения жидкости.
Большое значение имеет и сохранение достаточного кровоснабжения мозгового вещества почки. При серповидно-клеточной анемии нарушение концентрационной способности почки связано с тем, что патологически измененные эритроциты, входящие с током крови в прямые сосуды мозгового вещества, резко изменяют свою форму, соприкасаясь с гипертонической средой. Это уменьшает кровоток в узких сосудах мозгового слоя; ишемия только этой зоны снижает в ней накопление натрия, хлора, мочевины, в конечном счете нарушается функция концентрирования мочи. После обменного переливания крови концентрационная способность почки у этих больных временно восстанавливается до нормы. Другим примером может служить питрессинрезистентный ноктуральный нефрогенный несахарный диабет, описанный при пароксизмальной гипертензии. Он обусловлен усилением кровотока по прямым сосудам мозгового вещества; увеличение кровотока вызывает вымывание осмотически активных веществ из мозгового вещества почки в период пароксизмальной гипертензии, что сопровождается снижением осмоляльности мочи.
Процесс осмотического концентрирования мочи представляет собой совокупную работу всех составляющих почку элементов. Поэтому для эффективного концентрирования важное значение имеют количество функционирующих нефронов, тесная связь их друг с другом, способствующая деятельности противоточной системы, нормальные размеры отдельных сегментов нефронов. Снижение концентрационной способности при хронической почечной недостаточности (ХПН), в частности, зависит от резкого уменьшения количества функционирующих нефронов, увеличения расстояния между отдельными нефронами. Любые факторы, нарушающие соотношение между структурами мозгового слоя почки, приводят к снижению процесса осмотического концентрирования. Это может наблюдаться, например, при поликистозе или поражении мозгового вещества амилоидом.
Значительное снижение реабсорбции жидкости в проксимальном канальце вызывает усиление потока жидкости по петле Генле и вымывание из нее осмотически активных веществ. Это наблюдается при врожденном дефекте нефрона — укороченном проксимальном канальце, осмотическом диурезе любой этиологии (сахарный диабет, ХПН, вливание маннита и других осмотических диуретиков). Активный транспорт солей клетками толстого восходящего отдела петли Генле нарушается при гипокалиемии из-за ухудшения работы ионных насосов. Гиперкальциемия и избыток кальция в клетке снижают накопление натрия в мозговом веществе, угнетая его активный транспорт.
Увеличение проницаемости для воды стенки собирательных трубок инициируется АДГ. Из русла крови он проникает в межклеточную жидкость, достигает внешней поверхности плазматических мембран базальной и латеральной поверхности клетки. В этих мембранах находятся У2-рецепторы, АДГ взаимодействует с ними, и при участии G-белков активируется аденилатциклаза, способствующая преобразованию АТФ в цАМФ. Последний перемещается в область апикальной части клетки, и вследствие стимуляции протеинкиназы и ряда биохимических реакций осуществляется перестройка люминальной плазматической мембраны, увеличивается ее проницаемость для воды, в нее встраиваются водные каналы — аквапорины 2. Сказанное не отражает известной уже в настоящее время сложности внутриклеточных биохимических и ультраструктурных преобразований, наступающих после активации У2-рецептора, однако позволяет понять возможные причины нарушения осморегулирующей функции почки при некоторых видах фармакотерапии, нарушении ионного состава внутренней среды организма, эндокринопатиях.
Нарушение концентрационной способности почки наблюдается при гипокортицизме, гипопитуитаризме и гипотиреоидизме. Изменение ионного состава внутренней среды оказывает влияние на реакцию клеток собирательных трубок на АДГ. Взаимодействие гормона с клеткой ослабевает при гиперкальциемии, дефицит калия в организме сопровождается нарушением хода внутриклеточных процессов, активируемых АДГ и происходящих после образования цАМФ. Различные фармакологические средства, применяемые в клинике, могут вмешиваться в биохимические процессы, участвующие в осмотическом концентрировании. Среди них можно упомянуть анальгетики, некоторые диуретики, винкристин. Оказалось, что в увеличении проницаемости люминальной мембраны участвует цитоскелет, поэтому фармакологические средства, влияющие на состояние цитоскелета, сборку микрофиламентов и микротрубочек, изменяют реакцию клеток на АДГ.
Существенное значение в механизме действия этого гормона имеет его способность активировать образование ряда простагландинов в мозговом веществе почки. Их функциональное назначение состоит в снижении чувствительности клеток к АДГ, уменьшении реакции на АДГ. Казалось бы, складывается парадоксальная ситуация — один и тот же гормон увеличивает проницаемость собирательных трубок для воды и способствует секреции веществ, угнетающих собственную реакцию. Этот факт можно объяснить тем, что АДГ является основным (а возможно, и единственным) регулятором водного обмена, поэтому была необходимость создать систему снижения реакции на этот гормон. В этом и заключается двойственность его эффекта: он увеличивает реабсорбцию воды и способствует выработке вещества, снижающего чувствительность рецепторов к вазопрессину, тем самым противодействующего возрастанию осмотической проницаемости. Гормон быстро разрушается, и пока в кровь поступают все новые количества АДГ, сохраняется увеличенная реабсорбция воды, однако снижение секреции этого гормона быстро приводит к восстановлению водонепроницаемости стенки канальца. Влияние на эффект АДГ могут оказывать и лекарственные препараты, влияющие на синтез и инактивацию простагландинов.
Данные об увеличении АДГ проницаемости канальцев для мочевины позволяют понять давно известный в клинике факт, что очищение от мочевины зависит от величины диуреза — оно меньше при низком диурезе и значительно возрастает при увеличении мочеотделения, достигая 65 % от одновременно измеряемого очищения от креатинина или инулина при водном диурезе. Малый уровень мочеотделения обычно обусловлен значительной концентрацией в крови АДГ и, следовательно, большей реабсорбцией в канальцах воды и мочевины; обратные процессы наблюдаются при большой скорости диуреза, когда нет АДГ. Нельзя не учитывать и рациона, в котором должно быть достаточное количество не только солей, но и белка. При потреблении малобелковой пищи образуется недостаточное количество мочевины и вследствие этого не создается необходимого уровня ее накопления в мозговом слое почки. Все сказанное должно помочь в воссоздании «целостного образа» процесса осмотического концентрирования и лечь в основу последующего анализа причин, вызывающих нарушение одной из основных функций почки, связанных с ее участием в водно-солевом обмене и важных в клинике для характеристики функционального состояния почки.
источник
Способностью к осмотическому разведению мочи, т. е. способностью к выделению мочи с меньшей концентрацией осмотически активных веществ, а следовательно, с меньшим осмотическим давлением, чем плазма крови, обладают почки млекопитающих, птиц, рептилий, амфибий, пресноводных рыб и кругло-ротых. В то же время способностью к образованию мочи с большей концентрацией осмотически активных веществ, т. е. с большей осмотической концентрацией, чем кровь, обладают лишь почки теплокровных животных. Многие исследователи пытались разгадать физиологический механизм этого процесса, но лишь в начале 50-х годов XX века была обоснована гипотеза, согласно которой образование осмотически концентрированной мочи обусловлено деятельностью повороти о-п ротивоточной множительной системы в почке.
Принцип противоточного обмена достаточно широко распространен в природе и используется в технике. Механизм работы такой системы рассмотрим на примере кровеносных сосудов в конечностях арктических животных. Во избежание больших потерь тепла кровь в параллельно расположенных артериях и венах конечностей течет таким образом, что теплая артериальная кровь согревает охлажденную венозную кровь, движущуюся к сердцу (рис. 12.8, А). В стопу притекает артериальная кровь низкой температуры, что резко уменьшает теплоотдачу. Здесь такая система функционирует только как противоточный обменник; в почке же она обладает множительным эффектом, т. е. увеличением эффекта,
достигаемого в каждом из отдельных сегментов системы. Для лучшего понимания ее работы рассмотрим систему, состоящую из трех параллельно расположенных трубок (рис. 12.8, Б). Трубки I и II дугообразно соединены на одном из концов. Стенка, общая для обеих трубок, обладает способностью переносить ионы, но не пропускать воду. Когда в такую систему через вход I наливают раствор концентрации 300 мосмоль/л (рис. 12.8, Б, а) и он не течет, то через некоторое время в результате транспорта ионов в трубке I раствор станет гипотоническим, а в трубке II — гипертоническим. В том случае, когда жидкость течет по трубкам непрерывно, начинается концентрирование осмотически активных веществ (рис. 12.8, Б, б). Перепад их концентраций на каждом уровне трубки вследствие одиночного эффекта транспорта ионов не превышает 200 мосмоль/л, однако по длине трубки происходит умножение одиночных эффектов, и система начинает работать как противоточная множительная. Так как по ходу движения жидкости из нее извлекаются не только ионы, но и некоторое количество воды, концентрация раствора все более повышается по мере приближения к изгибу петли. В отличие от трубок I и II в трубке III регулируется проницаемость стенок для воды: когда стенка становится водопроницаемой — начинает пропускать воду, объем жидкости в ней уменьшается. При этом вода идет в сторону большей осмотической концентрации в жидкость возле трубки, а соли остаются внутри трубки. В результате этого растет концентрация ионов в трубке III и уменьшается объем содержащейся в ней жидкости. Концентрация в ней веществ будет зависеть от ряда условий, в том числе от работы противоточной множительной системы трубок I и II. Как будет ясно из последующего изложения, работа почечных канальцев в процессе осмотического концентрирования мочи похожа на описанную модель.
В зависимости от состояния водного баланса организма почки выделяют гипотоническую (осмотическое разведение) или, напротив, осмотически концентрированную (осмотическое концентрирование) мочу. В процессе осмотического концентрирования мочи в почке принимают участие все отделы канальцев, сосуды мозгового вещества, интерстициальная ткань, которые функционируют как поворотно-противоточная множительная система. Из 100 мл фильтрата, образовавшегося в клубочках, около 60— 70 мл ( 2 /з) реабсорбируется к концу проксимального сегмента. Концентрация осмотически активных веществ в оставшейся в канальцах жидкости такая же, как и в ультрафильтрате плазмы крови, хотя состав жидкости отличается от состава ультрафильтрата вследствие реабсорбции ряда веществ вместе с водой в проксимальном канальце (рис. 12.9). Далее канальцевая жидкость переходит из коркового вещества почки в мозговое, перемещаясь по петле нефрона до вершины мозгового вещества (где каналец изгибается на 180°), переходит в восходящий отдел петли и движется в направлении от мозгового к корковому веществу почки.
Функциональное значение различных отделов петли нефрона
неоднозначно. Поступающая из проксимального канальца, в тонкий нисходящий отдел петли нефрона жидкость попадает в зону почки, в интерстициальной ткани которой концентрация осмотически активных веществ выше, чем в корковом веществе почки. Это повышение осмоляльной концентрации в наружной зоне мозгового вещества обусловлено деятельностью толстого восходящего отдела петли нефрона. Его стенка непроницаема для воды, а клетки транспортируют С1
, Na + в интерстициальную ткань. Стенка нисходящего отдела петли проницаема для воды. Вода всасывается из просвета канальца в окружающую интерстициальную ткань по осмотическому градиенту, а осмотически активные вещества остаются в просвете канальца. Концентрация осмотически активных веществ в жидкости, поступающей из восходящего отдела петли в начальные отделы дистального извитого канальца, составляет уже около 200 мосмоль/кг НгО, т. е. она ниже, чем в ультрафильтрате. Поступление С1
и Na + в интерстициальную ткань мозгового вещества увеличивает концентрацию осмотически активных веществ (осмоляльную концентрацию) межклеточной жидкости в этой зоне почки. На такую же величину растет и осмоляль-ная концентрация жидкости, находящейся в просвете нисходящего
отдела петли. Это обусловлено тем, что через водопроницаемую стенку нисходящего отдела петли нефрона в интерстициальную ткань по осмотическому градиенту переходит вода, в то же время осмотически активные вещества остаются в просвете этого канальца.
Чем дальше от коркового вещества по длиннику почечного сосочка находится жидкость в нисходящем колене петли, тем выше ее осмоляльная концентрация. Таким образом, в каждых соседних участках нисходящего отдела петли имеется лишь небольшое нарастание осмотического давления, но вдоль мозгового вещества почки осмоляльная концентрация жидкости в просвете канальца и в интерстициальной ткани постепенно растет от 300 до 1450 мосмоль/кг Н2О.
На вершине мозгового вещества почки осмоляльная концентрация жидкости в петле нефрона возрастает в несколько раз, а ее объем уменьшается. При дальнейшем движении жидкости по восходящему отделу петли нефрона, особенно в толстом восходящем отделе петли, продолжается реабсорбция С1
и Na + , вода же остается в просвете канальца. В начальные отделы дисталь-ного извитого канальца всегда — и при водном диурезе, и при антидиурезе — поступает гипотоническая жидкость, концентрация осмотически активных веществ в которой менее 200 мосмоль/кг
При уменьшении мочеотделения (анти диурезе), вызванном инъекцией АДГ или секрецией АДГ нейрогипофизом при дефиците воды в организме, увеличивается проницаемость стенки конечных частей дистального сегмента (связующий каналец) и собирательных трубок для воды. Из гипотонической жидкости, находящейся в связующем канальце и собирательной трубке коркового вещества почки, вода реабсорбируется по осмотическому градиенту, осмоляльная концентрация жидкости в этом отделе увеличивается до 300 мосмоль/кг Н2О, т. е. становится изоосмотичной крови в системном кровотоке и межклеточной жидкости коркового вещества почки. Концентрирование мочи продолжается в собирательных трубках; они проходят параллельно канальцам петли нефрона через мозговое вещество почки. Как отмечалось выше, в мозговом веществе почки постепенно возрастает осмоляльная концентрация жидкости и из мочи, находящейся в собирательных трубках, реабсорбируется вода; концентрация осмотически активных веществ в жидкости просвета канальца выравнивается с таковой в интерстициальной жидкости на вершине мозгового вещества. В условиях дефицита воды в организме усиливается секреция АДГ, что увеличивает проницаемость стенок конечных частей дистального сегмента и собирательных трубок для воды.
В отличие от наружной зоны мозгового вещества почки, где повышение осмолярной концентрации основано главным образом на транспорте Na + и С1
, во внутреннем мозговом веществе почки это повышение обусловлено участием ряда веществ, среди которых важнейшее значение имеет мочевина — для нее стенки прок-
симального канальца проницаемы. В проксимальном канальце реабсорбируется до 50 % профильтровавшейся мочевины, однако в начале дистального канальца количество мочевины несколько больше, чем количество мочевины, поступившей с фильтратом. Оказалось, что имеется система внутрипочечного кругооборота мочевины, которая участвует в осмотическом концентрировании мочи. При антидиурезе АДГ увеличивает проницаемость собирательных трубок мозгового вещества почки не только для воды, но и для мочевины. В просвете собирательных трубок вследствие реабсорбции воды повышается концентрация мочевины. Когда проницаемость канальцевой стенки для мочевины увеличивается, она диффундирует в мозговое вещество почки. Мочевина проникает в просвет прямого сосуда и тонкого отдела петли нефрона. Поднимаясь по направлению к корковому веществу почки по прямому сосуду, мочевина непрерывно участвует в противоточном обмене, диффундирует в нисходящий отдел прямого сосуда и нисходящую часть петли нефрона. Постоянное поступление во внутреннее мозговое вещество мочевины, С1
и Na + , реабсорбируемых клетками тонкого восходящего отдела петли нефрона и собирательных трубок, удержание этих веществ благодаря деятельности противоточной системы прямых сосудов и петель нефрона обеспечивают повышение концентрации осмотически активных веществ во внеклеточной жидкости во внутреннем мозговом веществе почки. Вслед за увеличением осмоляльной концентрации окружающей собирательную трубку интерстициальной жидкости возрастает реабсорбция воды из нее и повышается эффективность осморегу-лирующей функции почки. Эти данные об изменении проницаемости канальцевой стенки для мочевины позволяют понять, почему очищение от мочевины уменьшается при снижении мочеотделения.
Прямые сосуды мозгового вещества почки, подобно канальцам петли нефрона, образуют противоточную систему. Благодаря такому расположению прямых сосудов обеспечивается эффективное кровоснабжение мозгового вещества почки, но не происходит вымывания из крови осмотически активных веществ, поскольку при прохождении крови по прямым сосудам наблюдаются такие же изменения ее осмотической концентрации, как и в тонком нисходящем отделе петли нефрона. При движении крови по направлению к вершине мозгового вещества концентрация осмотически активных веществ в ней постепенно возрастает, а во время обратного движения крови к корковому веществу соли и другие вещества, диффундирующие через сосудистую стенку, переходят в интер-стициальную ткань. Тем самым сохраняется градиент концентрации осмотически активных веществ внутри почки и прямые сосуды функционируют как противоточная система. Скорость движения крови по прямым сосудам определяет количество удаляемых из мозгового вещества солей и мочевины и отток реабсорбируемой воды.
В случае водного диуреза функции почек отличаются от описанной ранее картины. Проксимальная реабсорбция не изменяется,
в дистальный сегмент нефрона поступает такое же количество жидкости, как и при антидиурезе. Осмоляльность мозгового вещества почки при водном диурезе в три раза меньше, чем на максимуме антидиуреза, а осмотическая концентрация жидкости, поступающей в дистальный сегмент нефрона, такая же — приблизительно 200 мосмоль/кг НгО. При водном диурезе стенка конечных отделов почечных канальцев остается водопроницаемой, а из протекающей мочи клетки продолжают реабсорбировать Na + . В итоге выделяется гипотоническая моча, концентрация осмотически активных веществ в которой может снижаться до 50 мосмоль/кг НгО. Проницаемость канальцев для мочевины низкая, поэтому мочевина экскретируется с мочой, не накапливаясь в мозговом веществе почки.
Таким образом, деятельность петли нефрона, конечных частей дистального сегмента и собирательных трубок обеспечивает способность почек вырабатывать большие объемы разведенной (гипотонической) мочи — до 900 мл/ч, а при дефиците воды экскре-тировать всего 10—12 мл/ч мочи, в 4,5 раза более осмотически концентрированной, чем кровь. Способность почки осмотически концентрировать мочу исключительно развита у некоторых пустынных грызунов, что позволяет им длительное время обходиться без воды.
Дата добавления: 2015-06-04 ; Просмотров: 518 ; Нарушение авторских прав? ;
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
источник
Способность почек к образованию гипертоничной мочи обусловлена двумя факторами:
1) в мозговом веществе почек создается гипертоничная среда;
2) проницаемость собирательных трубочек для воды может существенно повышаться, и при этом осмолярность канальцевой жидкости становится такой же, как и гипертоничной среды мозгового вещества. Гипертоничная среда в мозговом веществе почек создается благодаря работе поворотно-противоточной системы. Деятельность этой системы обусловлена следующими основными факторами: 1) шпилькообразным изгибом петли Генле, благодаря которому жидкость в ее нисходящей и восходящей части, а также в параллельно расположенной собирательной трубочке течет в противоположных (противоточных) направлениях;
2) активным транспортом NaCl в толстом сегменте восходящей части петли Генле;
3) особенностями проницаемости разных отделов нефрона для воды и растворенных веществ.
В толстом сегменте восходящей части петли Генле активно реабсорбируется Na+ (это единственный требующий затрат энергии процесс в механизме концентрирования мочи). Поскольку этот сегмент непроницаем для воды, оттекающая из него жидкость становится гипото-ничной (с осмолярностью 50 мосм/л), а в интерстиции, напротив, создается гипертоничная среда. Нисходящая часть петли Генле, в которую из проксимального канальца поступает изотоничная жидкость, напротив, проницаема для воды, но непроницаема для NaCl и мочевины; в результате вода выходит из нее в гипертоничный интерстиции.
Продвигаясь по направлению к изгибу петли Генле, жидкость нисходящей части все больше теряет воду, становится все более гипертоничной (у взрослых ее осмолярность в области изгиба петли Генле достигает 1200 мосм/л), и концентрация Na+ и мочевины в ней нарастает. После перехода в тонкий сегмент восходящей части эпителий петли Генле становится непроницаемым для воды, но высоко проницаемым для Na+ и мочевины; в результате Na+ выходит из него в интерстиций по концентрационному градиенту. Мочевина, напротив, входит в тонкий сегмент, так как ее концентрация в интерстиций выше, чем в жидкости этого сегмента.
Это связано с тем, что мозговой (но не корковый) отдел собирательной трубочки проницаем для мочевины и эта проницаемость еще более возрастает под действием АДГ; поскольку же из-за ре-абсорбции воды концентрация мочевины в собирательной трубочке высока, это вещество выходит в интерстиций, еще более повышая его гипертоничность. Таким образом, мочевина претерпевает кругооборот, переходя из собирательной трубочки в интерстиций, а затем в восходящую часть петли Генле, и при этом вносит важный вклад в создание в интерстиций высоко гипертоничной среды.
Окончательная концентрация мочи зависит от присутствия АДГ. Этот гормон секретируется в ответ на увеличение осмолярности плазмы или уменьшение эффективного ОЦК. В отсутствие АДГ собирательная трубочка непроницаема для воды и из нее оттекает такая же ги-поосмолярная жидкость, как и поступающая в нее из дистального извитого канальца. Под действием АДГ в апикальную мембрану клеток собирательной трубочки внедряются водные каналы, образованные белком аквапорином-2; проницаемость собирательной трубочки для воды увеличивается, вода выходит в гипертоничный интерстиций мозгового вещества, и моча концентрируется.
Максимальная осмолярность мочи у взрослых — 1200 мосм/л, а у доношенного новорожденного — лишь 600—800 мосм/л. Концентрационная способность почек становится такой же, как у взрослых, к 12— 18 мес. Из-за этого у новорожденных легче развиваются дегидратация и гипернатриемия.
В ответ на увеличение осмолярности плазмы или снижение ОЦК сывороточная концентрация АДГ у новорожденного возрастает до таких же значений, как у взрослых. В то же время у новорожденных способность почки концентрировать мочу под действием АДГ снижена вследствие целого ряда факторов. Максимальная концентрация мочи зависит от гипертоничности мозгового вещества, которая у новорожденных ограничена из-за относительно малого потребления белков и образования мочевины, а также из-за меньшей реабсорбции NaCl в толстом сегменте восходящей части петли Генле. Почечные сосочки у новорожденных короче, что также снижает эффективность работы поворотно-противоточной системы.
Из-за увеличенного объема интерстиция и слабого развития прямых артериол диффузия мочевины и воды в почках новорожденных затруднена. Наконец, у новорожденных АДГ в меньшей степени повышает проницаемость собирательных трубочек для воды.
Женское молоко гипотонично. Новорожденные способны выводить этот избыток воды, поддерживая осмолярность крови на уровне 290 мосм/л; при этом осмолярность мочи у них, как и у взрослых, может снижаться до 50 мосм/л. Тем не менее способность к выведению свободной воды у новорожденных ограничена и потребление больших количеств гипотоничной жидкости может привести к гипонатриемии. Напротив, у взрослых с нормальной способностью почек к разведению мочи возможности выведения свободной воды огромны и поэтому гипонатриемия при потреблении даже очень больших объемов воды развивается редко.
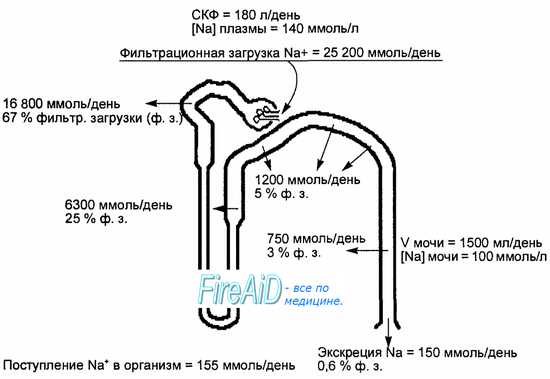 Почечные процессы, обеспечивающие поддержание баланса натрия.
Почечные процессы, обеспечивающие поддержание баланса натрия.
Прямые стрелки и цифры — реабсорбция и экскреция натрия в разных отделах нефрона, СКФ — скорость клубочковой фильтрации. При поступлени в сутки 155 ммоль натрия в организм, фильтруется в мочу за сутки 25 200 ммоль, из которых реабсорбируется 25 050 ммоль, в том числе 2/3 (67 %) в проксимальном отделе канальцев, а 1/3 (33 %) — в дистальном отделе. В результате за сутки с мочой выделяется 150 ммоль натрия. Оставшиеся 5 ммоль от поступившего количества натрия экскретируются из организма с калом и потом.
источник
Способностью к осмотич разведению мочи, т. е. способностью к выделению мочи с меньшей конц-ей осмотически активных в-в, а след-но, с меньшим осмотич давлением, чем плазма крови, обладают почки млекопитающих, птиц, рептилий, амфибий, пресноводных рыб и круглоротых. В то же время способностью к образ-ю мочи с большей конц-ей осмотически активных в-в, т. е. с большей осмотич конц-ей, чем кровь, обладают лишь почки теплокровных жив-х. Обусловлено деят-ю поворотно-противоточной множительной системы в почке. В завис-ти от сост-я водного баланса почки выд-ют гипотонич-ю (осмотич-е разведение) или осмотически концетрир-ную (осмотич концентрир-е) мочу. В процессе осмотич-го концетр-я мочи в почке принимают участие все отделы канальцев, сосуды мозгового в-ва, интерстиц ткань, кот-е функционируют как поворотно-противоточная множительная с-ма (↑ конц-я и ↓ объем содержащейся в ней жидкости). При ↓ мочеотдел-я (антидиурез), вызванном инъекцией АДГ или секрецией АДГ нейрогипофизом при дефиците воды в орг-ме, ↑ прониц-ть стенки конечных частей дист сегмента и собират трубок для воды. Т.О., деят-ть петли нефрона, конечных частей дистального сегмента и собират трубок обеспечивает способность почек вырабатывать большие объемы разведенной мочи — до 900 мл/ч, а при дефиците воды экскретировать всего 10-12 мл/ч мочи, в 4,5 раза более осмотически концентрированной, чем кровь.
Гомеостатические ф-ции почек.
Для поддержания почками постоянства объема и состава внутр среды существуют специальные системы рефлекторной регуляции, включающие специфич рецепторы, афферентные пути и нервные центры, где происходит переработка информации. Команды к почке поступают по эфферентным нервам или гумор путем.
Роль почек в осмо- и волюморегуляции. Почки — основной орган осморегуляции. Они обеспеч-ют выделение избытка воды из орг-ма в виде гипотонич мочи при ↑ содержании воды (гипергидратация) или экономят воду и экскретируют мочу, гипертоническую по отношению к крови, при обезвоживании орг-ма (дегидратация).
При избытке воды в орг-ме ↓ конц-я растворенных осмотически активных в-в в крови и ↓ ее осмоляльность. Это ↓ активность центральных осморецепторов, расположенных в области супраоптич ядра гипоталамуса, а также периферических осморецепторов, имеющихся в печени, почке и др органах, что приводит к ↓ секреции АДГ нейрогипофизом и ↑ выделения воды почкой.
При обезвоживании орг-ма или введении в сосудистое русло гипертонич р-ра NaCl ↑ конц-я осмотически активных в-в в плазме крови, возб-ся осморецепторы, ↑ секреция АДГ, ↑ всасывание воды в канальцах, ↓мочеотделение и выдел-ся осмотически концентрированная моча. Помимо осморецепторов, секрецию АДГ стимулируют натрийрецепторы. При введении в область III желудочка мозга гипертонич р-ра NaCl наблюдался антидиурез, если же вводить в то же место гипертонич р-р сахарозы, то мочеотделение не ↓.
Помимо осмо- и натрийрецепторов, уровень секреции АДГ определяет активность волюморецепторов, воспринимающих изменение объема внутрисосудистой и внеклеточной ж-ти. Ведущее значение в регуляции секреции АДГ имеют рецепторы, кот-е реагируют на изменение напряжения сосудистой стенки в области низкого давления. Прежде всего это рецепторы левого предсердия. При ↑ кровенаполнения левого предсердия активир-ся волюморецепторы и ↓ секреция АДГ, что вызывает ↑ мочеотделения. Возб-е волюморецепторов приводит к ↑экскреции почкой не только воды, но и Na.
Регуляция реабсорбции и секреции ионов в почечных канальцах осущ-ся несколькими гормонами. Реабсорбция Na ↑ в конечных частях дистального сегмента нефрона и собират трубках под влиянием альдостерона. В ↑ выделения Na почкой участвует натрийуретический гормон. Секрецию К + в дистальном сегменте и собират трубках ↑ альдостерон. Инсулин ↓ выделение К + . При ↓ конц-и Са в крови паращитовидные железы выделяют паратгормон, кот-й способ-ет нормализации уровня Са в крови. При гиперкальциемии стимулируется выделение в кровь парафолликулярными клетками щитовидной железы кальцитониналкот-й способствует ↓ концентрации Са 2+ в плазме крови.
Роль почек в регуляции кислотно-основного состояния. Почки участвуют в поддержании постоянства конц-и Н + в крови, экскретируя кислые продукты обмена. Активная р-я мочи у чел-ка и жив-х резко меняется в завис-ти от состояния кислотно-основного состояния орг-ма.
Это способствует участию почек в стабилизации рН плазмы крови на уровне 7,36. Мех-м подкисления мочи основан на секреции клетками канальцев Н + . Секретируемый Н + может связываться в просвете канальца с NH3, образуется ион аммония: NH3 + Н + =NH4+. Этот процесс способствует сбережению в орг-ме Na + и К + , кот-е реабсорб-ся в канальцах. Т.О., общая экскреция к-т почкой складыв-ся из 3 компонентов — титруемых к-т, аммония и гидрокарбоната. При питании мясом образ-ся большое кол-во к-т и моча становится кислой, а при потреблении растит пищи рН сдвиг-ся в основную сторону. При интенсивной физ работе из мышц в кровь поступает значит кол-во молочной и фосфорной к-т и почки ↑ выделение «кислых» продуктов с мочой.
Дата добавления: 2019-02-12 ; просмотров: 69 ; ЗАКАЗАТЬ РАБОТУ
источник
В почках предусмотрен механизм, позволяющий регулировать осмотическое давление плазмы крови за счет концентрирования или, наоборот, разведения мочи. Этот механизм получил название — поворотно-противоточно-множительный механизм, который локализован в петле Генле и собирательных трубках. Именно в этой части нефрона происходит изменение осмотической концентрации, или осмотического давления мочи, благодаря чему осуществляется регулирование объема выводимой жидкости и ее осмотического давления. Принцип работы этого механизма заключается в следующем: когда первичная моча проходит по собирательной трубке и направляется в чашечку, а затем в лоханку, из нее выходит вода, благодаря чему резко уменьшается объем конечной мочи, и возрастает концентрация в моче осмотически активных веществ. Если бы этот процесс не происходил, то ежедневно, помимо обычных 1—1,5 л мочи, выделялось еще 20—24 л мочи. Когда первичная моча входит в начальную часть собирательной трубки, ее осмотическое давление составляет примерно 290—300 мосмоль (милиосмоль — это единица концентрации осмотически активных веществ, или единица осмотического давления, например, раствор 140 ммоль хлористого натрия представляет собой 140 мосмоль натрия + 140 мосмоль хлора = 280 мосмоль, а раствор, содержащий 140 ммоль хлористого натрия и 10 ммоль хлористого калия имеет осмолярность, равную 140 + 140 + 10 +10 = 300 мосмоль/л.). По мере прохождения мочи по собирательной трубке вода покидает трубку и поэтому осмотическое давление (осмолярность мочи) возрастает до 900—1200 мосмоль, т. е. концентрируется в 3—4 раза. Для такого концентрирования необходимо, чтобы в интерстиции по мере приближения трубки к сосочку (по направлению к чашечке) росло осмотическое давление. В этом случае на каждом «этаже» вода будет покидать трубку и моча будет становиться все более концентрированной.
Выход воды из собирательной трубки происходит за счет осмотического давления, которое создается вокруг собирательной трубки в интерстиции. Это давление обусловлено наличием в интерстиции осмотически активных веществ — ионов натрия, хлора, мочевины. При этом необходимым условием должно быть постепенное повышение концентрации осмотически активных веществ по направлению от наружного слоя к внутреннему слою мозговой части почки. Осмотически активные вещества — натрий и хлор — появляются в интерстиции благодаря работе эпителия восходящей части петли Генле, за счет активного транспорта происходит выход ионов натрия и хлора из первичной мочи, идущей по восходящей части петли Генле. Мочевина появляется в интерстиции благодаря выходу из собирательных трубок за счет градиента концентрации между мочой, находящейся в собирательной трубке, и содержимого интерстиции (см. выше, реабсорбция мочевины). Для того, чтобы из восходящей части петли Генле происходила реабсорбция натрия и ионов хлора, в нисходящей части петли Генле осуществляется выход воды в интерстициальное пространство, где ионы натрия и хлора, вышедшие из восходящей части петли Генле создают осмотически активную среду. Рассмотрим пример. В нисходящую часть петли Генле входит первичная моча, осмолярность которой равна 300 мосмоль/л. По мере продвижения мочи к повороту за счет осмотически активной среды в интерстиции вода из этой порции мочи
будет уходить, и постепенно концентрация осмотически активных веществ будет возрастать — 400,500,600 мосмоль, достигая на вершине петли Генле максимальной концентрации, например, 1200 мосмоль. Затем, поднимаясь по восходящей части петли Генле, эпителий которой непроницаем для воды, но проницаем для ионов натрия и хлора, натрий и хлор за счет разности концентрации будут выходить в интерстиций (это и необходимо для конечной концентрации мочи в собирательных трубках), поэтому осмолярность первичной мочи по мере подъема ее по восходящей части петли Генле будет снижаться: 1000,900,800,700 мосмоль и т. д., а на вершине восходящей части петли Генле осмолярность может стать ниже исходной (например, 200,100 мосмоль). Когда моча попадает в собирательную трубку, то за счет высокой проницаемости для воды (но не для натрия и хлора), вода будет покидать собирательную трубку, а осмотическая концентрация (осмолярность) будет возрастать на тем большую величину, чем дальше проходит моча.
Итак, восходящая часть петли Генле создает условия для концентрации мочи как в нисходящей части петли Генле, так и в собирательных трубках.
Проницаемость собирательной трубки для воды зависит от уровня в крови вазопрессина (антидиуретического гормона): чем он выше, тем выше проницаемость для воды, тем меньше организм теряет воды, тем концентрированнее моча. И наоборот, когда уровень АДГ в крови снижается, тогда проницаемость для воды падает, и объем выделяемой мочи возрастает, а концентрация осмотически активных веществ в моче снижается.
Работа такого механизма позволяет организму тонко регулировать осмотическое давление плазмы крови и других жидкостей — при увеличении осмотического давления крови увеличивается реабсорбция воды в собирательных трубках, возрастает объем циркулирующей крови, что приводит к нормализации осмотического давления плазмы. Наоборот, при уменьшении осмотического давления почки в большем объеме, чем обычно, выделяют воду, что приводит к нормализации осмотического давления.
Важная роль в процессах концентрирования мочи принадлежит мозговому кровотоку. Если этот кровоток очень интенсивный, то из интерстиция удаляются осмотически активные вещества — натрий, хлор, мочевина. Тем самым снижается способность концентрировать мочу, возрастает диурез. Вот почему при повышении системного артериального давления вследствие роста интенсивности мозгового кровотока снижается концентрация осмотически активных веществ в мозговом слое почки и повышается диурез.
Для того, чтобы сохранить градиент концентрации осмотически активных веществ в мозговом слое (по направлению к внутренней части мозгового слоя он возрастает), капилляры мозгового слоя почки располагаются в виде петель, параллельно петлям Генле.
Итак, представленные данные позволяют понять принцип работы поворотно-противо-точно-множительной системы: за счет параллельного расположения трубок (нисходящей и восходящей частей петли Генле, собирательной трубки) создается противоток, в результате которого умножаются градиенты концентрации осмотически активных веществ.
источник
Способностью к образованию мочи с большей осмотической концентрацией, чем кровь, обладают лишь почки теплокровных животных. Многие исследователи пытались разгадать физиологический механизм этого процесса, но лишь в начале 50-х годов XX века была обоснована гипотеза, согласно которой образование осмотически концентрированной мочи связано с механизмом противоточно-поворотной множительной системы некоторых участков нефрона.
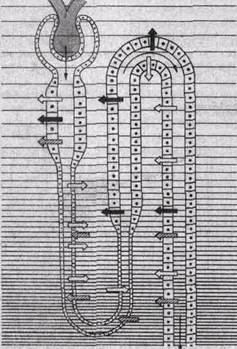
Принцип противоточного обмена достаточно широко распространен в природе и используется в технике. Механизм работы такой системы рассмотрим на примере кровеносных сосудов в конечностях арктических животных. Во избежание больших потерь тепла кровь в параллельно расположенных артериях и венах конечностей течет таким образом, что теплая артериальная кровь согревает охлажденную венозную кровь, движущуюся к сердцу (рис. 204). В стопу притекает артериальная кровь с низкой температурой, что резко уменьшает теплоотдачу. Здесь такая система функционирует только как противоточный обменник: в почке же она обладает множительным эффектом. Для лучшего понимания ее работы рассмотрим систему, состоящую из трех параллельно расположенных трубок. Трубки I и II дугообразно соединены на одном из концов (рис. 204, Б). Стенка, общая для обеих трубок, обладает способностью переносить соли, но она не пропускает воду. Когда в такую систему через вход 1 наливают жидкость с концентрацией 300 мосмоль/л и она не течет, то через некоторое время в результате транспорта солей в трубке I жидкость станет гипотонической, а в трубке 11 — гипертонической. В том случае, когда жидкость течет по трубкам непрерывно, начинается кон-центрирование солей. На каждом горизонтальном уровне перепад их концентраций вследствие одиночного эффекта транспорта солей не может превышать 200 мосмоль/л, однако по длине трубки происходит умножение одиночных эффектов и система начинает работать как противоточная множительная система. Так как по ходу движения жидкости из нее извлекаются не только соль, но и некоторое количество воды, концентрация раствора все более повышается по мере приближения к изгибу петли. В трубке III регу-
| Рис.204. Схема функционирования противоточно-поворотной системы. А — теплообменник в сосудистой системе конечностей арктических животных; обмен тепла между артериальной и венозной кровью способствует сбережению тепла и на каждом уровне не превышает 1° — 2° С. Б — модель противоточной множительной системы в начальном состоянии (а) и в период эффективного концентрирования мочи (б). Объяснение в тексте. |
 |
| ^— Активный транспорт Nа В зависимости от состояния водного баланса организма почки выделяют разведенную или концентрированную мочу. В процессе осмотического концентрирования мочи в почке принимают участие все отделы канальцев, сосуды мозгового вещества, интер-стициальная ткань. Из 100 мл фильтрата, образовавшегося в клубочках, ^з его реаб-сорбируются к концу проксимального сегмента. Оставшаяся в канальцах жидкость содержит осмотически активные вещества в такой же концентрации, как и ультрафильтрат плазмы крови, хотя и отличается от него по составу вследствие реабсорбции ряда веществ в предшествующих частях нефрона. Далее канальцевая жидкость переходит из коркового слоя почки в мозговое вещество — в нисходящий (тонкий) отдел петли нефрона (петля Генле) и движется до вершины почечного сосочка, где каналец изгибается на 180°, и моча переходит в восходящий отдел петли, расположенный параллельно ее нисходящему отделу. Функциональное значение различных отделов петли неоднозначно. Когда жидкость из проксимального отдела канальца поступает в тонкий нисходящий отдел петли нефрона, она попадает в зону почки, в интерстициальной ткани которой концентрация осмотически активных веществ выше, чем в коре почки. Это повышение осмолярной концентрации в наружной зоне мозгового вещества обусловлено деятельностью толстого восходящего отдела петли нефрона. Его стенка непроницаема для воды, а клетки транспортируют ионы С1 и Nа + в интерстициальную ткань. Стенка нисходящего отдела петли проницаема для воды, и поэтому вода всасывается из просвета канальца в окружающую межуточную ткань почки по осмотическому градиенту, а осмотически активные вещества остаются в просвете этого отдела канальца. Чем дальше от коры по продольной оси находится жидкость в нисходящем колене петли, тем выше ее осмолярная концентрация. В каждых соседних участках нисходящего отдела петли имеется лишь небольшое нарастание осмотического давления, но по длине петли осмолярная концентрация постепенно растет от 300 мосмоль/л почти до 1450 мосмоль/л. Иначе говоря, на вершине петли нефрона осмолярная концентрация жидкости возрастает в несколько раз и при этом объем ее уменьшается. При дальнейшем передвижении жидкости по восходящему отделу петли нефрона происходит реабсор-бция ионов С1 и Nа + , вода остается в просвете канальца, поэтому в начальные части дистального извитого канальца всегда поступает гипотоническая жидкость, концентрация осмотически активных веществ в которой менее 200 мосмоль/л. Из гипотонической жидкости по осмотическому градиенту реабсорбируется вода, осмолярная концентрация жидкости в этом отделе увеличивается, т. е. жидкость в просвете канальца становится изоосмотической. Окончательное концентрирование мочи происходит в собирательных трубках; они расположены параллельно канальцам петли нефрона, в мозговом веществе почки. Как отмечалось выше, в интерстициальной жидкости мозгового вещества почки возрастает осмолярная концентрация. Вследствие этого из жидкости собирательных трубок реабсорбируется вода и концентрация мочи в них увеличивается, уравновешиваясь со все повышающейся осмолярной концентрацией внутреннего мозгового вещества почки. В конечном счете выделяется гиперосмотическая моча, в которой максимальная концентрация осмотически активных веществ может быть равна осмолярной концентрации интерстициальной жидкости на вершине почечного сосочка (рис. 205). В условиях дефицита воды в организме усиливается секреция антидиуретического гормона гипофиза (АДГ), что увеличивает проницаемость стенок конечных частей дистального сегмента и собирательных трубок для воды. 14 -Физиология человека417 В отличие от наружной зоны мозгового вещества почки, где повышение осмолярности основано главным образом на транспорте хлоридов, увеличение осмолярной концентрации во внутренней зоне мозгового вещества почки зависит от нескольких механизмов. Особую роль в осмотическом концентрировании играет накопление мочевины. Стенки проксимального канальца проницаемы для мочевины. В этом отделе нефрона реабсорбируется до 50% профильтровавшейся мочевины. Однако при извлечении жидкости из извитого дистального канальца оказалось, что содержание мочевины даже несколько превышает ее количество, поступившее с фильтратом, и составляет около 1 !0%. Было показано, что имеется система внутрипочечного кругооборота мочевины, которая участвует в осмотическом концентрировании мочи. В просвете собирательных трубок вследствие реаб-сорбции воды повышается концентрация мочевины, АДГ увеличивает проницаемость собирательных трубок в мозговом веществе не только для воды, но и для мочевины. Когда увеличивается проницаемость канальцевой стенки для мочевины, она диффундирует в мозговое вещество почки. Постоянное поступление во внутреннее мозговое вещество мочевины, ионов С1 и Nа + , реабсорбируемых клетками тонкого восходящего отдела петли нефрона и собирательных трубок, обеспечивает повышение осмотической концентрации в мозговом веществе почки. Вслед за увеличением осмолярности окружающей собирательные трубки межуточной ткани возрастает и реабсорбция воды из них и повышается эффективность осморегулирующей функции почки. Изменение проницаемости канальцевой стенки для мочевины позволяет понять, почему очищение от мочевины уменьшается при снижении мочеотделения. Прямые кровеносные сосуды мозгового вещества почки, подобно канальцам петли нефрона, также образуют противоточную систему, играющую очень важную роль в осмотическом концентрировании. Благодаря особенностям расположения прямых сосудов обеспечивается эффективное кровоснабжение мозгового вещества почки, но не происходит вымывания осмотически активных веществ, поскольку в крови прямых сосудов наблюдаются такие же изменения осмотической концентрации, как и в тонком нисходящем отделе петли нефрона. При движении крови в ней постепенно возрастает осмотическая концентрация, а во время ее обратного движения к коре почки соли и другие растворенные вещества, диффундирующие через сосудистую стенку, переходят в интерстици-альную ткань. Тем самым сохраняется градиент концентрации осмотически активных веществ, т. е. прямые сосуды функционируют как противоточная система. Скорость движения крови по прямым сосудам влияет на количество удаляемых из мозгового вещества ионов Ма^ С1 и мочевины, участвующих в создании осмотического градиента, и отток реабсорбируемой воды. При водной нагрузке относительная проксимальная реабсорбция ионов и воды не изменяется, и в дистальный отдел нефрона поступает такое же количество жидкости, как и без нагрузки. При этом стенка дистальных отделов почечных канальцев остается водонепроницаемой, а из протекающей мочи клетки продолжают реабсорбировать соли натрия; при этом выделяется гипотоническая моча, концентрация осмотически активных веществ в которой ниже 50 мосмоль/л. Проницаемость канальцев для мочевины низкая, и она экскретируется с мочой, не накапливаясь в мозговом веществе почки. Собирательные трубки также обеспечивают реабсорбцию натрия, хлора и других ионов. Их основная функциональная особенность состоит в том, что реабсорбция веществ происходит в небольших количествах, но против наиболее значительного градиента, что обусловливает существенные различия концентрации ряда неорганических веществ в моче по сравнению с кровью. , Таким образом, деятельность петли нефрона, конечных частей дистального отдела собирательных трубок обусловливает способность почек человека при водной нарузке выделять большие объемы (до 900 мл/ч) разведенной, гипотонической мочи, а при дефиците воды в организме экскретировать мочи всего 10—12 мл/ч, в 4’/2 раза осмотически более концентрированной, чем кровь. Способность почки осмотически концентрировать мочу исключительно развита у некоторых пустынных грызунов, что позволяет им длительное время не пить воду. ГОМЕОСТАТИЧЕСКАЯ ФУНКЦИЯ ПОЧЕК Для поддержания почками постоянства объема и состава внутренней среды и прежде всего крови существуют специальные системы рефлекторной регуляции, включающие специфические рецепторы, афферентные пути и нервные центры, где происходит Рис.206. Схема регуляции водо- и электролитовыделительной функции почки. переработка информации. Команды к почке поступают по эфферентным нервам или гуморальным путем. В целом перестройка работы почки, ее приспособление к непрестанно изменяющимся условиям определяется преимущественно влиянием на гломеру-лярный и канальцевый аппарат различных гормонов: АДГ, альдостерона, паратгормона и многих других (рис. 206) Роль почек в осморегуляции и волюморегуляции. Почки являются основным органом осморегуляции. Они обеспечивают выделение избытка воды из организма в виде гипотонической мочи при увеличенном содержании воды (гипергидратация) или экономят воду и экскретируют мочу, гипертоническую по отношению к плазме крови при обезвоживании организма (дегидратация). При избыточном содержании воды в организме концентрация растворенных осмотически активных веществ в крови снижается и ее осмотическое давление падает. Это уменьшает активность центральных осморецепторов, расположенных в области супраоптического ядра гипоталамуса, а также периферических осморецепторов, имеющихся в печени, почке, селезенке и ряде других органов, что снижает выделение АДГ из нейрогипофиза в кровь и приводит к усилению выделения воды почкой. При обезвоживании организма (или введении в сосудистое русло гипертонического раствора хлорида натрия) увеличивается концентрация осмотически активных веществ в плазме крови, возбуждаются осморецепторы, усиливается секреция АДГ, возрастает факультативная реабсорбция воды, уменьшается мочеотделение и выделяется осмотически концентрированная моча. В эксперименте на животных было показано, что секреция АДГ возрастает при раздражении не только осморецепторов, но и специфических натриорецепторов. Поэтому после введения в область III желудочка мозга гипертонического раствора хлорида натрия наступает антидиурез, а после введения в ту же область гипертонических растворов сахара угнетения мочеотделения не наблюдается. Осморецепторы чутко реагируют на сдвиги концентрации осмотически активных веществ в плазме крови. При увеличении осмолярности плазмы крови на 1% концентрация’АДГ возрастает на 1 пг/мл (пикограмм равен 1 миллионной доле микрограмма). Переход же к состоянию максимального осмотического концентрирования мочи требует всего лишь 10-кратного возрастания количества АДГ в крови. Помимо информации от осморецепторов и натриорецепторов, уровень секреции АДГ зависит от активности волюморецепторов, реагирующих на изменение объема внутрисосудистой и внеклеточной жидкости. Ведущее значение в регуляции секреции АДГ имеют те волюморецепторы, которые реагируют на изменение напряжения сосудистой стенки в области низкого давления. В первую очередь следует назвать рецепторы левого предсердия, импульсы от которых передаются в ЦНС по афферентным волокнам блуждающего нерва. Изолированная активация волюморецепторов, возникающая в ответ на увеличение объема внутрисосудистой жидкости, приводит к повышению экскреции солей натрия и воды. Одновременная активация волюм- и осморецепторов в основном усиливает выведение воды, уменьшая ее реабсорбцию. Роль почек в регуляции ионного состава крови. Почки являются важнейшим эффек-торным органом в системе ионного гомеостаза. Новейшие данные свидетельствуют о существовании в организме систем регуляции баланса каждого из ионов. Для некоторых из ионов уже описаны специфические рецепторы, например натриорецепторы. Появились первые данные о рефлекторной регуляции баланса других ионов. Известны гормоны, регулирующие реабсорбцию и секрецию ионов в почечных канальцах. Реабсорбция натрия возрастает в конечных частях дистального отдела нефрона и собирательных трубках под влиянием гормона коры надпочечника альдосте-рона. Этот гормон выделяется в кровь при снижении концентрации натрия в плазме и уменьшении объема циркулирующей крови. Усиленное выделение натрия почкой вследствие угнетения его реабсорбции достигается при секреции в кровь так называемого натрийуретического гормона; его выработка возрастает при увеличении объема циркулирующей крови и повышении объема внеклеточной жидкости в организме. Хотя мысль о существовании этого гормона была высказана в конце 50-х годов, его структура и место секреции до сих пор не установлены. Альдостерон усиливает выделение калия с мочой. Уменьшает выделение калия ин-сулин. Экскреция калия тесно связана с кислотно-щелочным состоянием. Алкалоз сопровождается усилением выделения калия с мочой, а ацидоз — его уменьшением. При снижении концентрации кальция в крови паращитовидные железы выделяют паратгормон, который способствует нормализации уровня кальция в крови за счет увеличения его реабсорбции в почечных канальцах и повышения резсорбции кальция из костей. При гиперкальциемии под влиянием гастрина или подобного ему вещества, вырабатываемого в желудочно-кишечном тракте, стимулируется выделение в кровь клетками щитовидной железы гормона тирокальцитонина, который снижает концентрацию кальция в крови, способствует увеличению его экскреции почкой и переходу ионов Са’ 1 » 1 ‘ в кости. В почечных канальцах регулируется также уровень реабсорбции магния, хлора, сульфатов и других ионов. Роль почек в регуляции кислотно-основного состояния. Почки играют важную роль в поддержании постоянства концентрации ионов водорода в крови. Активная реакция мочи у человека и животных может очень резко изменяться, концентрации ионов водорода в моче при крайних состояниях работы почек различаются почти в 1000 раз (рН в некоторых случаях снижается до 4,5 или возрастает до 8,0, что и обеспечивает участие почек в стабилизации рН плазмы крови на уровне 7,36). Механизм закисления мочи основан на секреции клетками в просвет канальцев ионов водорода (рис. 207). В апикальной плазматической мембране и цитоплазме клеток различных отделов нефрона находится фермент карбоангидраза, катализирующий реакцию гидратации двуокиси углерода. Фильтрующийся из плазмы крови в просвет канальцев МаНСОз взаимодействует с ионами водорода, секретированными клеткой, приводя к образованию СОг. В просвете канальца ионы Н’ 1 ‘ связываются не только с НСОз, но и с такими соединениями, как двузамещенный фосфат (МагНРС^) и некоторыми другими, в результате чего увеличивается экскреция титруемых кислот с мочой. Это способствует восстановлению резерва Рис.207. Схема основных реакций, обусловливающих участие почки в регуляции кислотно-основного равновесия. Объяснение в тексте. оснований в плазме крови. Наконец, секретируемые ионы Н’ 1 ‘ могут связываться в просвете канальца с аммиаком, выделяющимся в клетке из глутамина и аминокислот (аммониогенез) и диффундирующим через мембрану в просвет канальца, в результате чего образуется ион аммония: Таким образом, общая экскреция кислот почкой складывается из трех компонентов: выделения НаСОз; титруемых кислот и выделения аммония МН^ Кислотовыделительная функция почек во многом зависит от состояния кислотно-основного состояния в организме. При питании мясом образуется больше кислот и моча становится кислой, а при потреблении растительной пищи рН мочи сдвигается в щелочную сторону. При интенсивной физической работе из мышц в кровь поступают значительные количества молочной и фосфорной кислот. В этих условиях почки увеличивают выделение кислых продуктов. При гиповентиляции легких происходит задержка углекислого газа и снижается рН крови — развивается дыхательный ацидоз, при гипервентиляции уменьшается содержание углекислого газа в крови, растет рН крови — возникает состояние дыхательного алкалоза. Если в крови нарастает содержание ацетоуксусной и р-оксимасляной кислот, как это может наблюдаться при сахарном диабете, развивается состояние метаболического ацидоза. Рвота, сопровождающаяся потерей соляной кислоты, приводит к метаболическому алкалозу. Легкие наряду с почками участвуют в нормализации кислотно-основного состояния. При дыхательном ацидозе увеличивается экскреция Н» 1 » и реабсорбция НСОз^ Метаболический ацидоз компенсируется гипервентиляцией легких. В конечном счете почки, стабилизируя концентрацию ионов Н’ 1 ‘ в плазме крови, поддерживают рН на.уровне 7,36. Экскреторная функция почек. Почки играют ведущую роль в выделении из крови нелетучих конечных продуктов обмена и чужеродных веществ, попавших во внутреннюю среду организма. В процессе метаболизма белков и нуклеиновых кислот образуются различные продукты азотистого обмена. Основными азотсодержащими веществами мочи у человека являются мочевина, мочевая кислота, креатинин и др. Катаболизм пуриновых оснований в организме человека останавливается на уровне образования мочевой кислоты (в клетках некоторых животных имеется полный набор ферментов, обеспечивающих распад пуриновых оснований до углекислоты и аммиака). Мочевая кислота в почке человека фильтруется в клубочках, затем реабсорбируется в каяальцах, часть мочевой кислоты секретируется клетками в просвет нефрона. Обычно у человека экскретируемая фракция мочевой кислоты довольно низкая—9,8%, что указывает на реабсорбцию значительных ее количеств в канальцах. В последние десятилетия непрерывно возрастает интерес к изучению механизмов транспорта мочевой кислоты в почечных канальцах в связи с резко возросшей частотой заболевания подагрой, при которой нарушен обмен мочевой кислоты. Весь образующийся в течение суток креатинин, источником которого служит креа-тинфосфорная кислота, выделяется почками с мочой. Его суточная продукция и экскреция зависят не столько от потребления мяса с пищей, сколько от массы мышц тела. Креатинин, как и мочевина, свободно фильтруется в почечных клубочках, но с мочой выводится весь профильтровавшийся креатинин, в то время как мочевина частично реабсорбируется в канальцах. Помимо перечисленных имеется еще очень много разнообразных веществ, непрестанно удаляемых почкой из крови. О том, какие вещества удаляет или разрушает почка, можно судить при изучении состава крови у людей с удаленными почками. В их крови, помимо мочевины, креатинина, мочевой кислоты, накапливаются гормоны (глюкагон, паратгормон, гастрин), ферменты (рибонуклеаза, ренин), производные индола, глюку-роновая кислота и многие другие вещества. Необходимо помнить, что даже физиологически ценные вещества при их избытке в крови начинают экскретироваться почкой. Это относится как к неорганическим веществам, о которых шла речь выше при описании осмо-, волюм- и ионорегулирующей функции почек, так и органическим веществам, таким, как глюкоза, аминокислоты. Повышенная экскреция этих веществ может в условиях патологии наблюдаться и при нормальной их концентрации в крови, когда нарушена работа клеток, реабсорбирующих то или иное профильтровавшееся вещество из канальцевой жидкости в кровь. Инкреторная функция почек. В почке вырабатывается несколько физиологически активных веществ, позволяющих рассматривать ее как важный инкреторный орган. Гранулярные клетки юкстагломерулярного аппарата под влиянием ряда стимулов (при уменьшении артериального давления в почке, снижении содержания натрия в организме и при переходе из горизонтального в вертикальное положение) секретируют ренин. Ренин, синтезируемый в гранулярных клетках, представляет собой протеолитический фермент. В плазме крови он отщепляет от «г-глобулина (ангиотензиноген) физиологически неактивный пептид, состоящий из 10 аминокислот — ангиотензин I. Затем от анги-отензина I отщепляются две аминокислоты и он превращается в очень активное сосудосуживающее вещество — ангиотензин II. Помимо влияния на сосуды и тем самым на величину артериального давления, ангиотензин II, в частности, усиливает секрецию альдостерона, усиливает чувство жажды, регулирует реабсорбцию натрия в канальцах. Клетки почки извлекают из плазмы крови образующийся в печени прогормон — витамин Оз и превращают его в физиологически очень активный гормон. Этот стероид стимулирует образование кальцийсвязывающего белка в кишечнике, способствует освобождению кальция из костей, регулирует его реабсорбцию в почечных канальцах. В почке синтезируется активатор плазминогена — урокиназа. В мозговом веществе почки образуются простагландины, в том числе простагландин Аа, который называют также медуллином. Они участвуют в регуляции общего и почечного кровотока, вызывают натрийурез, уменьшают чувствительность клеток к АДГ. В почке образуется эри-трогенин, который способствует превращению в плазме крови неактивного эритропоэ-тиногена в эритропоэтин, стимулирующий эритропоэз в костном мозге. В почке вырабатывается брадикинин, являющийся сильным вазодилататором. Метаболическая функция почек. Важной стороной работы почек является их участие в обмене белков, углеводов и липидов. Не следует смешивать понятия метаболизма самих почек (т. е. процессы обмена веществ в почке, обеспечивающие все многообразие выполняемых ею функций) и метаболическую функцию почек, связанную с их участием в обеспечении постоянства содержания в крови ряда органических веществ. В почечных клубочках фильтруются низкомолекулярные белки, пептиды. Клетки проксимального ка-нальца нефрона расщепляют их до аминокислот, которые через базальную плазматическую мембрану транспортируются во внеклеточную жидкость и затем в кровь. Это способствует восстановлению в организме фонда аминокислот. При заболеваниях почки эта функция может нарушаться. В почке имеется система новообразования глюкозы — глюконеогенез. Так, при длительном голодании почки могут синтезировать половину от общего количества глюкозы, поступающей в кровь. В почке синтезируется фосфатидилинозит, являющийся необходимым компонентом плазматических мембран. Значение почки в липидном обмене состоит в том, что свободные жирные кислоты в ее ткани могут включаться в состав три-ацилглицерина и фосфолипидов и в виде этих соединений могут поступать в кровь. Дата добавления: 2016-03-27 ; просмотров: 1375 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ источник |